

Potentially Beneficial Effects of St. John’s Wort (Hypericum perforatum) in Patients with Metabolic Syndrome
Hideya Mizuno* ![]() , Ayako Taketomi
, Ayako Taketomi![]() , Toshikatsu Nakabayashi
, Toshikatsu Nakabayashi![]()
- School of Pharmacy and Pharmaceutical Sciences, Mukogawa Women’s University, 11-68 Koshien Kyuban-cho, Nishinomiya, Hyogo 663-8179, Japan
* Correspondence: Hideya Mizuno![]()
Academic Editor: Srinivas Nammi
Received: June 27, 2018 | Accepted: September 3, 2018 | Published:September 28, 2018
OBM Integrative and Complementary Medicine 2018, Volume 3, Issue 3 doi:10.21926/obm.icm.1803021
Special Issue: Herbal Medicines for the Treatment of Metabolic Syndrome
Recommended citation: Mizuno H, Taketomi A, Nakabayashi T. Potentially Beneficial Effects of St. John's Wort (Hypericum perforatum) in Patients with Metabolic Syndrome. OBM Integrative and Complementary Medicine 2018; 3(3): 021; doi:10.21926/obm.icm.1803021.
© 2018 by the authors. This is an open access article distributed under the conditions of the Creative Commons by Attribution License, which permits unrestricted use, distribution, and reproduction in any medium or format, provided the original work is correctly cited.
Abstract
St. John’s wort (SJW), or Hypericum perforatum, is a perennial herb that is used to treat depression in several countries. Based on the available evidence, it can be hypothesized that SJW would also have beneficial prophylactic and therapeutic effects in patients with metabolic syndrome. In this review, we summarize and discuss the best designed studies that have investigated the effects of SJW extract on adipocyte differentiation, the inflammatory process, and glucose metabolism, all of which are relevant to the development of metabolic syndrome. Despite some lack of clarity in reporting of research and discrepancies in interpretation, we consider that SJW is able to enhance differentiation of adipocytes via activation of peroxisome proliferator-activated receptor gamma and increase expression of adiponectin, exert anti-inflammatory effects by inhibiting expression of pro-inflammatory adipokines, and maintain normal glucose metabolism by protecting pancreatic beta cells and peripheral target tissues against changes in insulin secretion and insulin sensitivity. Although further research is necessary, there is encouraging evidence that SJW may help to attenuate the pathologic process associated with metabolic syndrome.
Keywords
St. John’s wort; hypericum perforatum; metabolic syndrome; adipocyte differentiation; adiponectin; peroxisome proliferator-activated receptor gamma; inflammation; insulin resistance
1. Introduction
Metabolic syndrome is a pathological disorder that is caused mainly by the accumulation of visceral fat, which increases the risk of type 2 diabetes and cardiovascular disease [1]. The increased volume of adipose tissue that occurs in obesity reflects an increase in the number and/or size of adipocytes [2]. The increase in size of adipocytes could be attributed to their excessive lipid content. Adipose tissue not only stores lipids but also functions as an endocrine organ that secretes hormones known as adipokines [3]. Enlarged adipocytes and activated infiltrating M1 macrophages that occur in obesity secrete pro-inflammatory adipokines, including tumor necrosis factor alpha (TNF-α), interleukin (IL)-6, and monocyte chemoattractant protein (MCP)-1 [4,5,6,7]. These pro-inflammatory adipokines cause disorders of glucose and lipid metabolism by impairing metabolically important tissues, such as adipose, muscle, and endocrine pancreatic tissue, contributing to the development of diabetes or cardiovascular disease [3]. Therefore, metabolic syndrome can be viewed as a state of chronic low-grade inflammation caused by pro-inflammatory adipokines. In contrast, normal adipocytes secrete metabolically important adipokines, such as adiponectin or leptin [8,9,10]. Adiponectin has a variety of roles in glucose and lipid metabolism, including regulation of glucose uptake, oxidation of fatty acids, sensitization of the liver and muscle to insulin, and anti-inflammatory effects [8,9,10].
St. John’s wort (SJW), or Hypericum perforatum, is a perennial herb that is used to treat depression in several countries [11,12]. Generally, SJW is not associated with severe side effects. Mild side effects, such as gastrointestinal upset, minor palpitations, photosensitivity, fatigue, restlessness, dry mouth, and headache occur occasionally [11]. It is well known that SJW is an inducer of cytochrome P450 and may reduce the actions of some drugs [11]. SJW contains various bioactive compounds, including naphthodianthrones (hypericin and pseudohypericin), phloroglucinols (hyperforin and adhyperforin), biflavonoids (biapigenin and amentoflavone), glycosides (rutin and quercetin), and other groups of flavonoids [13,14,15]. Hypericin and hyperforin are the main ingredients responsible for the antidepressant effects of SJW [14,15] and are contained in the flowers and leaves of the plant. The concentrations of hypericin and pseudohypercin in the extracts vary from 0.03% to 0.3% and those of hyperforin and adhyperforin vary from almost 0% to 6% [14,15]. The content of hyperforin and adhyperforin in SJW is strongly influenced by the concentration of alcohol in the extraction solvent [15].
The therapeutic effects of SJW and its ingredients have been investigated in many potential indications, including cancer [16,17,18,19,20,21] as well as cardiovascular [22,23], inflammatory, and allergic diseases [24,25,26,27]. However, although the effects of SJW on depression have been extensively studied, its effects on other diseases have not been elucidated. Various studies, including one by our research group, have studied the effects of SJW extract on the components of metabolic syndrome. Based on the findings of these studies, it can be hypothesized that SJW may also have a beneficial role in the prevention and treatment of metabolic syndrome. In this review, we summarize and discuss the more interesting studies that have investigated the effects of SJW extract on the differentiation of adipocytes, inflammation, and glucose metabolism, all of which are relevant to the development of metabolic syndrome.
2. Effects of SJW on Differentiation of Adipocytes and Secretion of Adiponectin
Differentiation of adipocytes is tightly regulated by transcription factors that include peroxisome proliferator-activated receptor gamma (PPARγ) and the CCAAT/enhancer-binding protein (C/EBP) family [28,29]. PPARγ is a member of the PPAR subfamily of nuclear hormone receptors [2,30] and plays a particularly important role in the promotion of adipogenesis. PPARγ also regulates the expression of adiponectin, which binds to the promoter region of the adiponectin gene and facilitates gene transcription [31]. Therefore, activation of PPARγ by agonists such as thiazolidinediones, which are used in the treatment of type 2 diabetes [32,33], increases the number of small adipocytes by promoting differentiation and decreases the number of large adipocytes by inducing apoptosis [34,35], thereby improving glucose and lipid metabolism by secreting adiponectin.
Several studies have investigated the effects of SJW extract on differentiation of adipocytes and secretion of adiponectin. Amini et al. reported that extracts from the flowers and leaves of SJW (but not from its roots) inhibited differentiation of 3T3-L1 preadipocytes [36]. The same research group also showed that an extract from the flowers of SJW inhibited insulin-sensitive glucose uptake in vitro [37]. These extracts were prepared with 80% ethanol extraction. Concentrations of extract in the range of 25–50 µg/mL were used for the experiments. Commercially available extracts from the flower tops of SJW have also been investigated [38]. In our study, treatment of 3T3-L1 cells with SJW extract facilitated differentiation of adipocytes without cell hypertrophy and increased the expression and secretion of adiponectin. There were discrepancies between these findings and those of similar in vitro studies. Notably, 50 µg/mL of SJW extract was used in our study [38] which was a slightly higher concentration than that used in previous in vitro reports [36,37], suggesting that the content and/or concentration of the bioactive ingredients in our SJW extract possibly differed from those used in other studies. The differences in the ingredients might be responsible for the varying study results. However, further research is necessary to identify the ingredients responsible for these differences in the in vitro experiments. Importantly, at almost the same time as we published our in vitro report, the same research group reported that administration of SJW extract to mice increased total adiponectin levels in white adipose tissue and insulin sensitivity, although the serum levels of adiponectin were not affected [39]. These results were in contrast with those of their in vitro studies [36,37]. They speculated that the metabolism and/or absorption of SJW extract in vivo might have led to the contradictory results in the in vitro studies, and concluded that SJW does not promote metabolic dysfunction in adipose tissue. Although there was some discordance in the results of similar in vitro studies, our in vitro results were consistent with the most recent in vivo results reported from the same group.
Furthermore, SJW extract increased the expression of PPARγ in our study [38]. Therefore, we expected that SJW extract would facilitate the differentiation of adipocytes and expression of adiponectin via activation of PPARγ. However, GW9662, a PPARγ antagonist, did not decrease the increased expression of adiponectin caused by treatment with SJW extract. Similarly, GW9662 did not inhibit the increased differentiation of adipocytes or expression of adiponectin induced by telmisartan, a partial PPARγ agonist [40]. A possible explanation for these negative findings is that the inhibitory effect of GW9662 on PPARγ does not continue during the differentiation period. There was a report suggesting that the half-life of GW9662 was shorter than that of a PPARγ agonist in a cultured cell model [41]. Therefore, it is assumed that one of the ingredients in SJW extract acts as a PPARγ agonist and facilitates differentiation of adipocytes and expression of adiponectin via activation of PPARγ. We also need to consider that some ingredients of SJW extract might act through a PPARγ-independent pathway. Further investigation is necessary to clarify this mechanism.
Following this, we wondered which ingredient in SJW would be responsible for this activity. A recent report indicated that grape seed-derived polyphenols decreased the size of adipocytes and increased the number of adipocytes overexpressing PPARγ in obese rats [42]. That report is consistent with the known effects of SJW extract [38] and PPARγ agonists [34,35]. SJW extract includes a variety of polyphenols that might induce differentiation of adipocytes and expression of adiponectin via activation of PPARγ. Further investigation of the effect of polyphenols in SJW extract on 3T3-L1 cell lines and in vivo studies are needed to confirm whether polyphenols are responsible for the activity of SJW.
3. Anti-inflammatory Effect of SJW
Expression of pro-inflammatory adipokines such as TNF-α, IL-6, and MCP-1 are secreted by enlarged adipocytes and/or infiltrating M1 macrophages; this expression is regulated by nuclear factor (NF)-κB, which is a critical inflammatory transcription factor [43,44]. NF-κB also regulates the expression of cyclooxygenase 2, which catalyzes the formation of prostaglandins, including prostaglandin E2 (PGE2), a pro-inflammatory lipid mediator from arachidonic acid [45]. The activity of NF-κB is regulated by endogenous κB inhibitors (IκB) that form complexes with NF-κB to keep it in an inactive state in the cytoplasm [46]. Phosphorylated IκBα is degraded by the ubiquitin proteasome system, resulting in release and nuclear translocation of NF-κB that then regulates gene expression [47]. Therefore, inhibition of NF-κB reduces the expression of pro-inflammatory cytokines and usually protects cells from apoptosis. However, in some types of cells, including cancer cells, inhibition of NF-κB leads cells to apoptosis because NF-κB becomes constitutively active and is associated with tumorigenesis via inhibition of apoptosis signaling. The NF-κB signaling pathway in inflammation and cancer has been reviewed in detail by Hoesel et al [48].
Previously, we have investigated the anti-inflammatory activity of SJW in adipocytes differentiated from 3T3-L1 preadipocytes [38]. TNF-α-induced upregulation of IL-6 and MCP-1 mRNA levels was significantly decreased in the presence of SJW extract. Furthermore, the SJW extract attenuated the TNF-α-induced increase in IκBα phosphorylation, indicating that SJW mediated inhibition of NF-κB activation.
In addition to our study in adipocytes, there is evidence showing that SJW has anti-inflammatory activity in a variety of other cells and tissues. SJW extract reduced paracetamol-induced inflammation in the livers of mice by inhibiting production of IL-1β, TNF-α, and interferon-γ [49]. Hammer et al. showed that pseudohypercin and hyperforin, which are major ingredients in SJW extract, inhibited production of PGE2 in RAW264.7 macrophage cells within the range of concentrations present in SJW extract [26]. Other ingredients in SJW, including rutin and quercetin, have anti-inflammatory effects, but exert these effects at higher concentrations than those present in SJW extract. Koeberle et al. also indicated that hyperforin suppressed production of PGE2 by inhibiting PGE2 synthase-1, which is the major PGE2 synthase present under pathologic conditions related to inflammation and cancer [50] in vivo [27]. Furthermore, hyperforin inhibited activation of NF-κB in various types of cells both in vivo and in vitro, affording effective protection against inflammatory damage or tumor growth [51,52,53]. Hyperforin also inhibited 5-lipoxygenase, which catalyzes the formation of pro-inflammatory lipid mediators known as leukotrienes in vitro and in vivo [24,25]. These reports suggest that hyperforin is the primary anti-inflammatory ingredient in SJW extract and might work alongside other ingredients.
4. Effect of SJW on Glucose Metabolism
Inflammation induced by enlarged adipocytes in skeletal muscle leads to the development of insulin resistance, which decreases the uptake of circulating glucose into cells and increases the blood glucose level [3]. This chronic inflammatory condition is considered to play an important role in impairment of glucose metabolism and the development of type 2 diabetes.
Several groups have studied the effects of SJW extract on glucose metabolism. Tian et al. performed glucose/insulin tolerance tests and measured the glucose infusion rate in insulin-resistant mice treated with SJW extract [54]. The test results improved significantly in the mice treated with the SJW extract, suggesting that SJW improves glucose metabolism. The same investigators also explored the molecular mechanism by which this improvement occurs. Protein tyrosine phosphatase (PTP)1B is a negative regulator of insulin signaling [55], and its inhibition is considered to improve insulin sensitivity. SJW extract inhibited P1PTB activity in vitro and reduced expression of PTP1B in skeletal muscle in a mouse model [54]. Significantly, the SJW preparation used in their study contained mainly naphthodianthrones (hypericin and pseudohypericin). Thus, the inhibitory effect on PTP1B should be attributed to these components. The study reported by Tian et al. suggests that SJW, and in particular its naphthodianthrone components, facilitates insulin-dependent glucose uptake in skeletal muscle [54]. The main two active ingredients of SJW extract appear to be complementary in inhibiting inflammatory signaling in pancreatic beta cells and improving insulin sensitivity in peripheral tissues. Furthermore, studies using isolated pancreatic islets or beta cell lines indicate that SJW extract and hyperforin inhibit pro-inflammatory cytokine-induced activation of STAT-1 and NF-κB, impairment of insulin secretion, and apoptosis [52,56,57]. Although there are few relevant studies as of yet, it is anticipated that SJW improves glucose metabolism by improving insulin sensitivity and inhibiting inflammatory signaling.
5. Conclusion
In this review, we have summarized the studies that have investigated the effects of SJW on differentiation of adipocytes, inflammation, and glucose metabolism. These studies indicate that SJW enhances differentiation of adipocytes via activation of PPARγ and increased expression of adiponectin, has anti-inflammatory effects by inhibiting expression and action of pro-inflammatory adipokines, and might contribute to maintain glucose homeostasis in pathologic conditions favoring development of insulin resistance and impaired insulin secretion. Figure 1 summarizes the effects of SJW in metabolically important tissues. Although further investigation is necessary to identify the active ingredient(s) in SJW and the mechanisms involved, there is evidence to suggest that SJW has the potential to attenuate the pathologic process associated with metabolic syndrome.
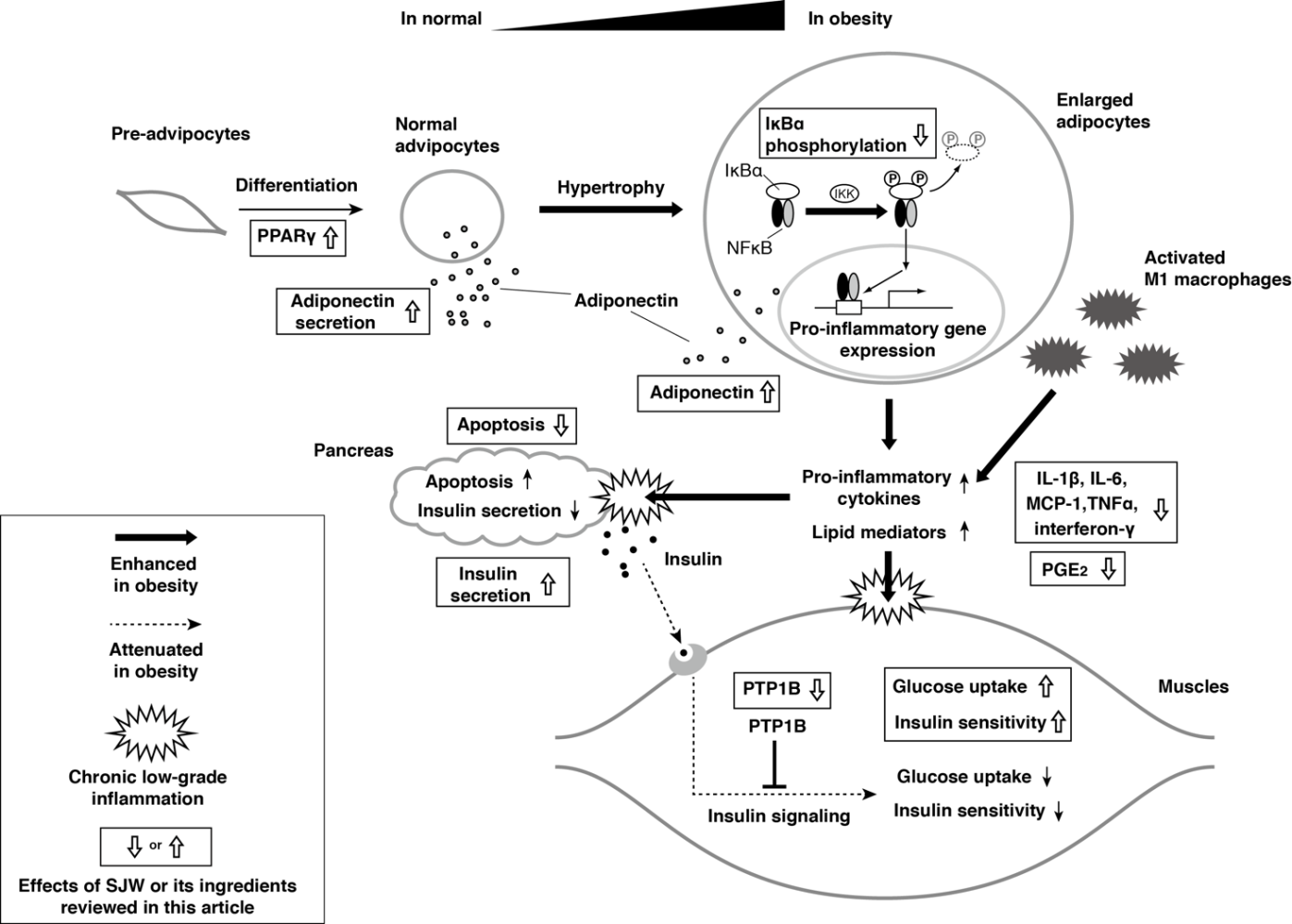
Figure 1 Schematic summary of the effects of SJW. IL, interleukin; MCP-1, monocyte chemoattractant protein-1; PPARγ, peroxisome proliferator-activated receptor gamma; PG, prostaglandin; PTP1B, protein tyrosine phosphatase 1B; SJW, St. John’s wort; TNF-α, tumor necrosis factor alpha.
Acknowledgments
We would like to thank Editage (www.editage.jp) for English language editing.
Author Contributions
Dr. Hideya Mizuno, Ms. Ayako Taketomi, and Dr. Toshikatsu Nakabayashi contributed to the writing and proofreading of this article.
Funding
None
Competing Interests
The authors have declared that no competing interests exist.
References
- Emanuela F, Grazia M, Marco DR, Maria Paola L, Giorgio F, Marco B. Inflammation as a link between obesity and metabolic syndrome. J Nutr Metab. 2012; 2012: 476380. [CrossRef] [Google scholar] [PubMed]
- Spiegelman BM, Flier JS. Adipogenesis and obesity: rounding out the big picture. Cell. 1996; 87: 377–389 [CrossRef] [Google scholar] [PubMed]
- Harwood HJ. The adipocyte as an endocrine organ in the regulation of metabolic homeostasis. Neuropharmacology. 2012; 63: 57–75. [CrossRef] [Google scholar] [PubMed]
- Christiansen T, Richelsen B, Bruun JM. Monocyte chemoattractant protein-1 is produced in isolated adipocytes, associated with adiposity and reduced after weight loss in morbid obese subjects. Int J Obes (Lond). 2005; 29: 146–150. [CrossRef] [Google scholar] [PubMed]
- Lau DCW, Dhillon B, Yan H, Szmitko PE, Verma S. Adipokines: molecular links between obesity and atherosclerosis. Am J Physiol Heart Circ Physiol. 2005; 288: H2031–2041. [CrossRef] [Google scholar] [PubMed]
- Sewter CP, Digby JE, Blows F, Prins J, O'Rahilly S. Regulation of tumour necrosis factor-alpha release from human adipose tissue in vitro. J Endocrinol. 1999; 163: 33–38. [CrossRef] [Google scholar] [PubMed]
- Hill AA, Reid Bolus W, Hasty AH. A decade of progress in adipose tissue macrophage biology. Immunol Rev. 2014; 262: 134–152. [CrossRef] [Google scholar] [PubMed]
- Scherer PE. Adipose tissue: from lipid storage compartment to endocrine organ. Diabetes. 2006; 55: 1537–1545. [CrossRef] [Google scholar] [PubMed]
- Matsuzawa Y. The metabolic syndrome and adipocytokines. FEBS Lett. 2006; 580: 2917–2921. [CrossRef] [Google scholar] [PubMed]
- Kadowaki T, Yamauchi T, Kubota N, Hara K, Ueki K, Tobe K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J Clin Invest. 2006; 116: 1784–1792. [CrossRef] [Google scholar] [PubMed]
- Lawvere S, Mahoney MC. St. John's wort. Am Fam Physician. 2005; 72: 2249–2254. [Google scholar]
- Solomon D, Adams J, Graves N. Economic evaluation of St. John's wort (Hypericum perforatum) for the treatment of mild to moderate depression. J Affect Disord. 2013; 148: 228–234. [CrossRef] [Google scholar] [PubMed]
- Wölfle U, Seelinger G, Schempp CM. Topical application of St. John's wort (Hypericum perforatum). Planta Med. 2014; 80: 109–120. [Google scholar]
- Barnes J, Anderson LA, Phillipson JD. St John's wort (Hypericum perforatum L.): a review of its chemistry, pharmacology and clinical properties. J Pharm Pharmacol. 2001; 53: 583–600. [CrossRef] [Google scholar] [PubMed]
- Linde K. St. John's wort - an overview. Forsch Komplementmed. 2009; 16: 146–155. [CrossRef] [Google scholar] [PubMed]
- Roscetti G, Franzese O, Comandini A, Bonmassar E. Cytotoxic activity of Hypericum perforatum L. on K562 erythroleukemic cells: differential effects between methanolic extract and hypericin. Phytother Res. 2004; 18: 66–72. [CrossRef] [Google scholar] [PubMed]
- Valletta E, Rinaldi A, Marini M, Franzese O, Roscetti G. Distinct Hypericum perforatum L. total extracts exert different antitumour activity on erythroleukemic K562 cells. Phytother Res. 2018; 376: 407. [CrossRef] [Google scholar] [PubMed]
- You MK, Kim HJ, Kook JH, Kim HA. St. John's wort regulates proliferation and apoptosis in MCF-7 human breast cancer cells by inhibiting AMPK/mTOR and activating the mitochondrial pathway. Int J Mol Sci. 2018; 19: 966. [CrossRef] [Google scholar] [PubMed]
- Hostanska K, Reichling J, Bommer S, Weber M, Saller R. Hyperforin a constituent of St John's wort (Hypericum perforatum L.) extract induces apoptosis by triggering activation of caspases and with hypericin synergistically exerts cytotoxicity towards human malignant cell lines. Eur J Pharm Biopharm. 2003; 56: 121–132. [CrossRef] [Google scholar] [PubMed]
- Lee JS, Lee MS, Oh WK, Sul JY. Fatty acid synthase inhibition by amentoflavone induces apoptosis and antiproliferation in human breast cancer cells. Biol Pharm Bull. 2009; 32: 1427–1432. [CrossRef] [Google scholar] [PubMed]
- Chen JH, Chen WL, Liu YC. Amentoflavone induces anti-angiogenic and anti-metastatic effects through suppression of NF-κB activation in MCF-7 cells. Anticancer Res. 2015; 35: 6685–6693. [Google scholar]
- Abd El Motteleb DM, Abd El Aleem DI. Renoprotective effect of Hypericum perforatum against diabetic nephropathy in rats: Insights in the underlying mechanisms. Clin Exp Pharmacol Physiol. 2017; 44: 509–521. [CrossRef] [Google scholar] [PubMed]
- Lin Y, Zhang JC, Fu J, Chen F, Wang J, Wu ZL, et al. Hyperforin attenuates brain damage induced by transient middle cerebral artery occlusion (MCAO) in rats via inhibition of TRPC6 channels degradation. J Cereb Blood Flow Metab. 2013; 33: 253–262. [CrossRef] [Google scholar] [PubMed]
- Albert D, Zündorf I, Dingermann T, Müller WE, Steinhilber D, Werz O. Hyperforin is a dual inhibitor of cyclooxygenase-1 and 5-lipoxygenase. Biochem Pharmacol. 2002; 64: 1767–1775. [CrossRef] [Google scholar] [PubMed]
- Feisst C, Pergola C, Rakonjac M, Rossi A, Koeberle A, Dodt G, et al. Hyperforin is a novel type of 5-lipoxygenase inhibitor with high efficacy in vivo. Cell Mol Life Sci. 2009; 66: 2759–2771. [CrossRef] [Google scholar] [PubMed]
- Hammer KDP, Hillwig ML, Solco AKS, Dixon PM, Delate K, Murphy PA, et al. Inhibition of prostaglandin E(2) production by anti-inflammatory hypericum perforatum extracts and constituents in RAW264.7 mouse macrophage cells. J Agric Food Chem. 2007; 55: 7323–7331. [CrossRef] [Google scholar] [PubMed]
- Koeberle A, Rossi A, Bauer J, Dehm F, Verotta L, Northoff H, et al. Hyperforin, an anti-inflammatory constituent from St. John's wort, inhibits microsomal prostaglandin E(2) synthase-1 and suppresses prostaglandin E(2) formation in vivo. Front Pharmacol. 2011; 2: 7. [CrossRef] [Google scholar] [PubMed]
- Siersbaek R, Nielsen R, Mandrup S. PPARgamma in adipocyte differentiation and metabolism--novel insights from genome-wide studies. FEBS Lett. 2010; 584: 3242–3249. [CrossRef] [Google scholar] [PubMed]
- Moseti D, Regassa A, Kim W-K. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int J Mol Sci. 2016; 17. doi: 10.3390/ijms17010124 [CrossRef] [Google scholar] [PubMed]
- Tontonoz P, Hu E, Spiegelman BM. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell. 1994; 79: 1147–1156. [CrossRef] [Google scholar] [PubMed]
- Iwaki M, Matsuda M, Maeda N, Funahashi T, Matsuzawa Y, Makishima M, et al. Induction of adiponectin, a fat-derived antidiabetic and antiatherogenic factor, by nuclear receptors. Diabetes. 2003; 52: 1655–1663. [CrossRef] [Google scholar] [PubMed]
- Lehmann JM, Moore LB, Smith-Oliver TA, Wilkison WO, Willson TM, Kliewer SA. An antidiabetic thiazolidinedione is a high affinity ligand for peroxisome proliferator-activated receptor gamma (PPAR gamma). J Biol Chem. 1995; 270: 12953–12956. [CrossRef] [Google scholar] [PubMed]
- Nolte RT, Wisely GB, Westin S, Cobb JE, Lambert MH, Kurokawa R, et al. Ligand binding and co-activator assembly of the peroxisome proliferator-activated receptor-gamma. Nature. 1998; 395: 137–143. [CrossRef] [Google scholar] [PubMed]
- Okuno A, Tamemoto H, Tobe K, Ueki K, Mori Y, Iwamoto K, et al. Troglitazone increases the number of small adipocytes without the change of white adipose tissue mass in obese Zucker rats. J Clin Invest. 1998; 101: 1354–1361. [CrossRef] [Google scholar] [PubMed]
- Xiao Y, Yuan T, Yao W, Liao K. 3T3-L1 adipocyte apoptosis induced by thiazolidinediones is peroxisome proliferator-activated receptor-gamma-dependent and mediated by the caspase-3-dependent apoptotic pathway. FEBS J. 2010; 277: 687–696. [CrossRef] [Google scholar] [PubMed]
- Amini Z, Boyd B, Doucet J, Ribnicky DM, Stephens JM. St. John's wort inhibits adipocyte differentiation and induces insulin resistance in adipocytes. Biochem Biophys Res Commun. 2009; 388: 146–149. [CrossRef] [Google scholar] [PubMed]
- Richard AJ, Amini ZJ, Ribnicky DM, Stephens JM. St. John's wort inhibits insulin signaling in murine and human adipocytes. Biochim Biophys Acta. 2012; 1822: 557–563. [CrossRef] [Google scholar] [PubMed]
- Hatano T, Sameshima Y, Kawabata M, Yamada S, Shinozuka K, Nakabayashi T, et al. St. John's wort promotes adipocyte differentiation and modulates NF-κB activation in 3T3-L1 cells. Biol Pharm Bull. 2014; 37: 1132–1138. [CrossRef] [Google scholar] [PubMed]
- Fuller S, Richard AJ, Ribnicky DM, Beyl R, Mynatt R, Stephens JM. St. John's wort has metabolically favorable effects on adipocytes in vivo. Evid Based Complement Alternat Med. 2014; 2014: 862575–862578. [CrossRef] [Google scholar] [PubMed]
- Shiota A, Shimabukuro M, Fukuda D, Soeki T, Sato H, Uematsu E, et al. Activation of AMPK-Sirt1 pathway by telmisartan in white adipose tissue: A possible link to anti-metabolic effects. Eur J Pharmacol. 2012; 692: 84–90. [CrossRef] [Google scholar] [PubMed]
- Li X, Ycaza J, Blumberg B. The environmental obesogen tributyltin chloride acts via peroxisome proliferator activated receptor gamma to induce adipogenesis in murine 3T3-L1 preadipocytes. J Steroid Biochem Mol Biol. 2011; 127: 9–15. [CrossRef] [Google scholar] [PubMed]
- Pascual-Serrano A, Arola-Arnal A, Suárez-García S, Bravo FI, Suárez M, Arola L, et al. Grape seed proanthocyanidin supplementation reduces adipocyte size and increases adipocyte number in obese rats. Int J Obes (Lond). 2017; 41: 1246–1255. [CrossRef] [Google scholar] [PubMed]
- Ajuwon KM, Spurlock ME. Adiponectin inhibits LPS-induced NF-kappaB activation and IL-6 production and increases PPARgamma2 expression in adipocytes. Am J Physiol Regul Integr Comp Physiol. 2005; 288: R1220–1225. [CrossRef] [Google scholar] [PubMed]
- Zoico E, Garbin U, Olioso D, Mazzali G, Fratta Pasini AM, Di Francesco V, et al. The effects of adiponectin on interleukin-6 and MCP-1 secretion in lipopolysaccharide-treated 3T3-L1 adipocytes: role of the NF-kappaB pathway. Int J Mol Med. 2009; 24: 847–851. [CrossRef] [Google scholar] [PubMed]
- Serhan CN, Savill J. Resolution of inflammation: the beginning programs the end. Nat Immunol. 2005; 6: 1191–1197. [CrossRef] [Google scholar] [PubMed]
- Baeuerle PA, Baltimore D. I kappa B: a specific inhibitor of the NF-kappa B transcription factor. Science. 1988; 242: 540–546. [CrossRef] [Google scholar] [PubMed]
- Chen ZJ, Parent L, Maniatis T. Site-specific phosphorylation of IkappaBalpha by a novel ubiquitination-dependent protein kinase activity. Cell. 1996; 84: 853–862. [CrossRef] [Google scholar] [PubMed]
- Hoesel B, Schmid JA. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer. BioMed Central; 2013; 12: 86. [Google scholar]
- Hohmann MSN, Cardoso RDR, Fattori V, Arakawa NS, Tomaz JC, Lopes NP, et al. Hypericum perforatum reduces paracetamol-induced hepatotoxicity and lethality in mice by modulating inflammation and oxidative stress. Phytother Res. 2015; 29: 1097–10101. [CrossRef] [Google scholar] [PubMed]
- Samuelsson B, Morgenstern R, Jakobsson P-J. Membrane prostaglandin E synthase-1: a novel therapeutic target. Pharmacol Rev. 2007; 59: 207–224. [CrossRef] [Google scholar] [PubMed]
- Lee S-K, Kim J-E, Kim Y-J, Kim M-J, Kang T-C. Hyperforin attenuates microglia activation and inhibits p65-Ser276 NFκB phosphorylation in the rat piriform cortex following status epilepticus. Neurosci Res. 2014; 85: 39–50. [CrossRef] [Google scholar] [PubMed]
- Novelli M, Menegazzi M, Beffy P, Porozov S, Gregorelli A, Giacopelli D, et al. St. John's wort extract and hyperforin inhibit multiple phosphorylation steps of cytokine signaling and prevent inflammatory and apoptotic gene induction in pancreatic β cells. Int J Biochem Cell Biol. 2016; 81: 92–104. [CrossRef] [Google scholar] [PubMed]
- Chen W-T, Chen Y-K, Lin S-S, Hsu F-T. Hyperforin suppresses tumor growth and NF-κB-mediated anti-apoptotic and invasive potential of non-small cell lung cancer. Anticancer Res. 2018; 38: 2161–2167. [Google scholar]
- Tian J-Y, Tao R-Y, Zhang X-L, Liu Q, He Y-B, Su Y-L, et al. Effect of Hypericum perforatum L. extract on insulin resistance and lipid metabolic disorder in high-fat-diet induced obese mice. Phytother Res. 2015; 29: 86–92. [CrossRef] [Google scholar] [PubMed]
- Johnson TO, Ermolieff J, Jirousek MR. Protein tyrosine phosphatase 1B inhibitors for diabetes. Nat Rev Drug Discov. 2002; 1: 696–709. [CrossRef] [Google scholar] [PubMed]
- Menegazzi M, Novelli M, Beffy P, D’Aleo V, Tedeschi E, Lupi R, et al. Protective effects of St. John's wort extract and its component hyperforin against cytokine-induced cytotoxicity in a pancreatic beta-cell line. Int J Biochem Cell Biol. 2008; 40: 1509–1521. [CrossRef] [Google scholar] [PubMed]
- Novelli M, Beffy P, Menegazzi M, De Tata V, Martino L, Sgarbossa A, et al. St. John's wort extract and hyperforin protect rat and human pancreatic islets against cytokine toxicity. Acta Diabetol. 2014; 51: 113–121. [CrossRef] [Google scholar] [PubMed]
