

Deciphering the Epigenetic Landscape of Suicidal Behaviour: A Review of Current Findings, Caveats and Future Directions
Stefania Policicchio *![]() , Emma L Dempster †
, Emma L Dempster †![]() , Therese M Murphy †
, Therese M Murphy †![]()
University of Exeter Medical School, Barrack Road, EX2 5DW, Exeter, UK
† These authors contributed equally to this work.
* Correspondence: Stefania Policicchio![]()
Received: June 20, 2018 | Accepted: September 10, 2018 | Published: October 06, 2018
OBM Genetics 2018, Volume 2, Issue 4 doi:10.21926/obm.genet.1804039
Academic Editors: Stéphane Viville and Marcel Mannens
Special Issue: Epigenetic Mechanisms in Health and Disease
Recommended citation: Policicchio S, Dempste EL, Murphy TM. Deciphering the Epigenetic Landscape of Suicidal Behaviour: A Review of Current Findings, Caveats and Future Directions. OBM Genetics 2018;2(4):039; doi:10.21926/obm.genet.1804039.
© 2018 by the authors. This is an open access article distributed under the conditions of the Creative Commons by Attribution License, which permits unrestricted use, distribution, and reproduction in any medium or format, provided the original work is correctly cited.
Abstract
Suicide is the second leading cause of death globally among young people and the tenth leading cause of death across all ages. Approximately 800,000 people die by suicide every year representing a significant global health burden. Despite this burden, the molecular pathology of suicide remains poorly understood. A number of recent studies have shown that epigenetic alterations are associated with suicidal behaviour. These epigenetic mechanisms, which act to regulate gene expression via modifications to DNA, histone proteins and chromatin, change with age and in response to specific environmental and psychosocial factors —providing a mechanism for the interaction between genotype and the environment. The present review briefly outlines the main epigenetic mechanisms involved in gene regulation and discusses recent findings of epigenetic alterations in suicidal behaviour, their caveats and the future direction of this emerging field of research.
Keywords
Epigenetics; suicide; DNA methylation; histone modification; non-coding RNA
1. Introduction
Suicide is a serious public health problem that can have lasting harmful effects on individuals, families, and communities [1]. Suicide is the second leading cause of death among 15–29 year olds worldwide and ranks amongst the 20th leading causes of death across all ages [2]. Approximately 800,000 people die by suicide every year [2] representing a significant global health burden. Suicidal behaviour (SB) spans a spectrum ranging from suicidal ideation (SI) to suicide attempt (SA) and completed suicide [3,4]. Up to one-third of individuals with SI have a SA within 1 year; individuals who have had a SA have 1.6% risk of suicide within the year [5]. Despite its economic and social burden, the underlying aetiology of SB remains poorly understood. Genetic epidemiological studies have found evidence of family clustering of SB and have highlighted a clear genetic susceptibility to SB [6,7] with heritability estimates ranging from 30-50% [8,9,10]. A significant proportion of the heritability for SB and SA appears to be independent of psychiatric diseases as when the heritability of SA is corrected for transmission of psychiatric disorders, specific heritability is between 17% and 36% [8,11,12,13,14,15].
Pre-existing knowledge of biological systems likely to be associated with SB, such as serotonergic neurotransmission and neurotrophic factors, resulted in several candidate-gene association studies [7,14,15,16,17] which have largely yielded inconsistent results. Similarly, large-scale genome-wide association studies (GWAS) [18,19,20] have failed to identify robust associations suggesting that the risk of SB is highly polygenic in nature and that individual gene variants are likely to account only for a very small proportion of the total phenotypic variability. Other factors, such as the environment, behavioural traits, lifestyle and coping mechanisms, are essential regulators of suicide risk and likely to account for more sizeable effects [21]. Recently, increased understanding of epigenetic processes that occur in the brain has opened promising avenues in suicide research. The epigenome is potentially malleable—changing with age [22] and in response to specific environmental [23] and psychosocial factors [24] —providing a mechanism for the interaction between genotype and the environment [25]. Epigenetic processes, including DNA methylation, have recently been implicated in the aetiology of numerous mental health disorders [26,27,28,29,30,31,32,33] and SB [34,35]. The present review will briefly introduce epigenetic mechanisms, focussing primarily on DNA methylation, histone modifications and non-coding RNAs and explore their relevance to suicide and SB, before discussing future directions and caveats of this emerging field of research and its potential to further our understanding of SB.
1.1 Beyond Genome-Wide Association Studies: a Role for Epigenetic Variation in Suicide and SB
The term epigenetics, literally meaning ‘above’ genetics, was originally coined to refer to the study of any potentially stable and heritable changes in gene expression or cellular phenotype that occurs without changes in Watson–Crick base-pairing of DNA [36]. However, the epigenome, which is the chemical structure surrounding our genome, can alter the function of our genes in response to the environment [37], and remains dynamic throughout life [37,38,39].
Epigenetic variation is regulated in a tissue-specific manner by stochastic, genetic and environmental factors [40]. In the brain, epigenetic variation is thought to influence the gene expression profile of neurons and glial cells [41,42]. Therefore it is possible that alterations of epigenetic profiles in the brain may induce long-term behavioural consequences. As such they have the potential to play a crucial role in determining predisposition to SB [43]. The most commonly studied epigenetic mechanisms that can influence gene expression are: molecular modifications of the DNA sequence, post-translational histone modifications and non-coding RNA gene silencing [44].
1.2 DNA Modifications
DNA methylation is the covalent modification at the C5 carbon of cytosine residues, occurring mainly at CG dinucleotides (CpG) (See Figure 1). In somatic cells, approximately 80% of CpGs are methylated [45]; the remaining unmethylated CpGs tend to be concentrated around gene promoters (CpG islands) [37]. A Recent study has reported 5-methyl cytosine (5mC) to be central to the establishment of tissue-specific gene expression, cell differentiation, genomic imprinting, and X-inactivation [46]. When found in promoter regions of genes, CpG methylation is generally associated with transcriptional repression or downregulation of RNA transcription [47]. However, DNA methylation has also been described in other regions of the genome and the transcriptional effect in those areas is not consistent [24]. Previous research has suggested a role for global DNA methylation as a potential marker of suicide risk. A significantly higher level of global methylation (5mC %) has been reported in peripheral blood of psychiatric patients with a history of SA when compared with psychiatric controls [35]. Similarly, an increase in DNA methylation in the brain region BA47 was found to be 8 times greater in the group who died by suicide relative to controls [44]. Taken together these findings, although requiring further replication, suggest that global DNA methylation may be a biomarker of suicide risk in psychiatric patients and support the idea that blood tissue may be reasonably considered a valid proxy to monitor brain changes.
Although DNA methylation has been considered as a stable epigenetic mark, studies in the past decade have revealed that this modification is not as static as once thought. Recently, research has provided evidence that 5mC can be oxidized to 5-hydroxymethylcytosine (5hmC) in a reaction catalysed by the ten–eleven translocation (TET) enzymes [48,49,50] along with two other oxidative products: 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). Growing evidence suggests that these demethylation states may play an important biological role [51,52].
1.3 Histone Modifications
Chromatin exists as two functional structures: euchromatin which is the ‘‘opened’’ state associated with gene transcription and heterochromatin which is the ‘‘closed’’ state associated with gene silencing. Proteins forming this complex structure are called histones (H2A, H2B, H3, and H4). These globular proteins, with a tail of amino-acids, can be covalently modified by the addition or the removal of chemical groups at specific residues on their N-terminus tails (See Figure 1) [37]. Histone modifications are able to alter the access of transcriptional machinery to the DNA by modifying the condensation of chromatin [37]. Although there are several molecular modifications histone acetylation/deacetylation and methylation are the two most commonly studied.
Studies examining the role of histone modifications in SB are limited to the study of candidate genes [53,54], thus genome-wide approaches have not yet been undertaken.
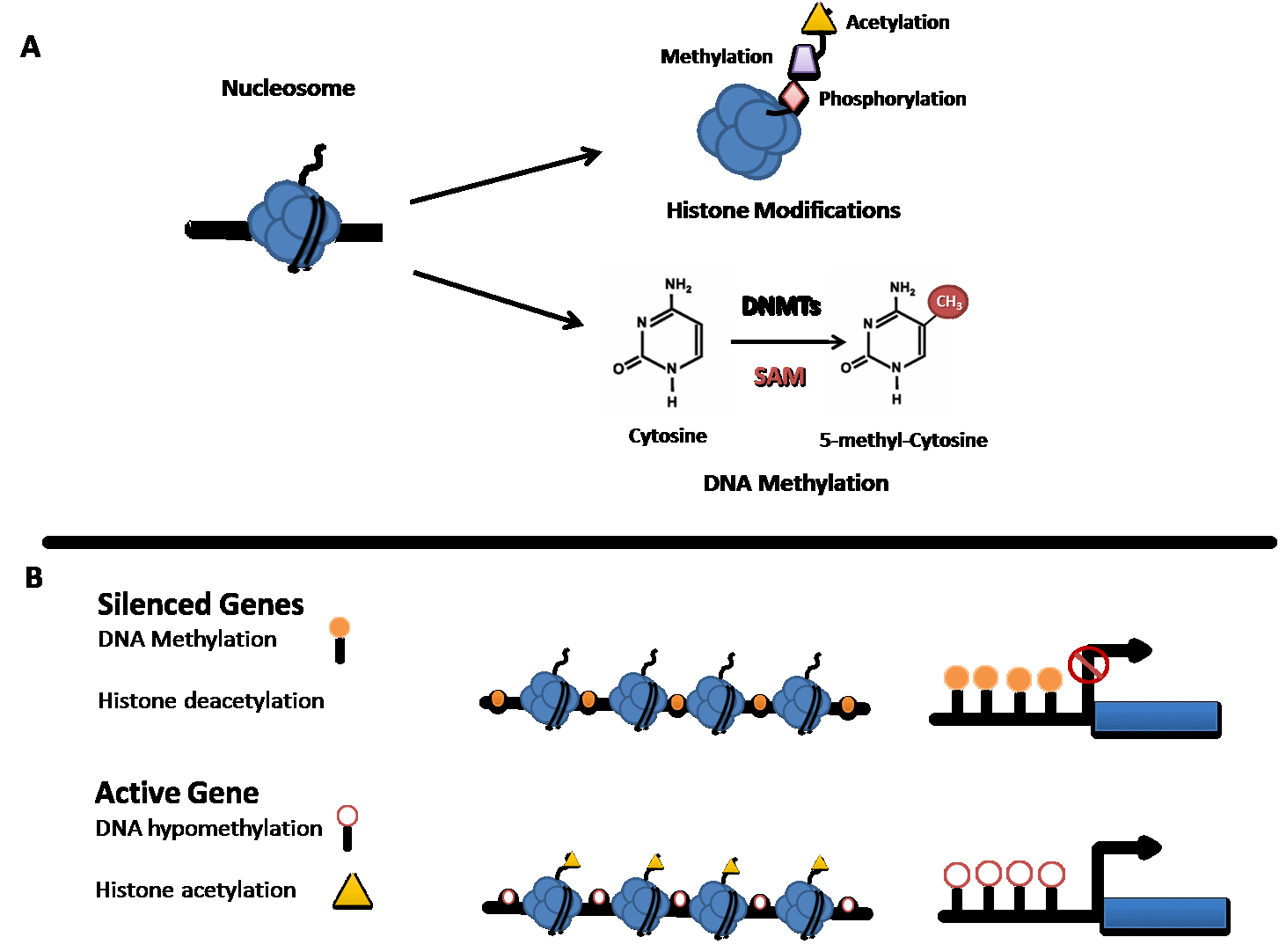
Figure 1 Epigenetic modifications and transcriptional regulation. (A) Figure illustrates common modifications that occur on the tails of histone proteins and the conversion of Cytosine to 5-methyl-Cytosine by a group of enzymes called DNA methyltransferases (DNMTs) and S-Adenosyl-L-methionine (SAM). (B) Transcriptionally inactive genes are generally characterised by DNA Methylation at CpG dinucleotides in the promoter region of genes and histone deacetylation. In contrast, transcriptionally active genes are generally characterised by absence of DNA methylation (DNA hypomethylation) and histone acetylation.
1.4 Non-Coding RNAs
Another mechanism of epigenetic regulation of gene expression is mediated by non-coding RNAs (ncRNAs), functional RNA molecules that are transcribed from DNA but not translated into proteins. These regulatory RNAs function to regulate gene expression at the transcriptional and post-transcriptional level. Several classes of ncRNAs exist, including microRNAs (miRNAs), short interfering RNAs (siRNAs), and piwi-interacting RNAs (piRNAs) also referred to as short ncRNAs (<30 nts) to distinguish them from the long non-coding RNAs (lncRNAs) (>200 nts). Both major groups are shown to play a role in heterochromatin formation, histone modification, DNA methylation targeting and gene silencing [55]. To date, miRNAs are the best characterised ncRNA group. They act as regulators of gene expression and protein translation in many tissues, including brain, by binding to the target mRNA and contributing to its degradation through the RNA-induced silencing complex (RISC) complex or by directly inhibiting translation [56]. MiRNAs are known to play a critical role in developmental processes, including neurodevelopment and synaptic plasticity and they have been implicated in the pathogenesis of various neuropsychiatric disorders like schizophrenia, major depression and bipolar disorders [55,56,57,58]. The specific dysregulation of miRNA function in suicide is just beginning to be appreciated, as recently reviewed [56]. Cataloguing the expression of miRNAs and additional small RNAs in healthy human brain as well as in major neuropsychiatric diseases and suicide may lead to new therapeutic targets and insight into disease aetiology [55].
LncRNAs represent another interesting class as they are enriched for expression in the brain [59] and are developmentally regulated [60] but less evolutionarily conserved than other RNA species. Although preclinical studies have begun to unravel how lncRNAs may contribute to emotional control [61], their role in SB is currently unknown.
2. Common Neurobiological Pathways Epigenetically Dysregulated in Suicide
In the last decade, understanding the role of epigenetic mechanisms in SB has implicated the role of key biological pathways, including hypothalamic pituitary adrenal (HPA) axis, stress response, polyamine system, neurotrophic signalling and lipid metabolism. A comprehensive overview of genes epigenetically dysregulated in suicide and SB is reported in Table 1 and Table 2. Common pathways frequently targeted by epigenetic and miRNA dysregulation are illustrated in Figure 2 and discussed in detail below.
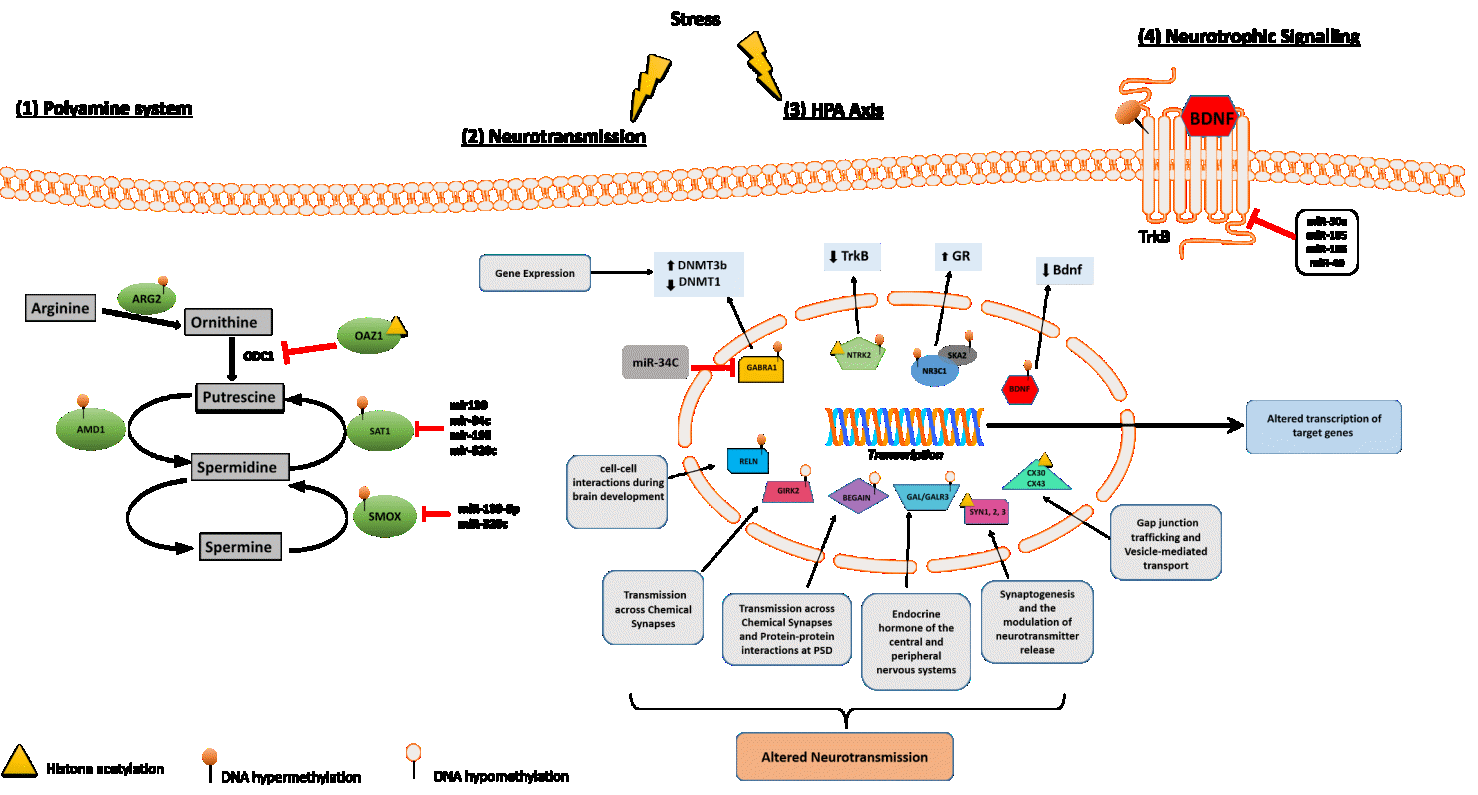
Figure 2 Molecular pathways involved in suicide behaviour. (1) Polyamine system: several enzymes involved in the regulation of polyamine intracellular concentration exhibit epigenetic regulatory mechanisms in suicide through DNA methylation (AMD1, ARG2), histone methylation (OAZ1) and/or post-translational mechanisms (SAT1, SMOX) as miRNAs regulation. (2) Neurotransmission: many genes known to play a key role at chemical and/or electrical synapses have been reported as hypomethylated (BEGAIN, GIRK2, GALR3), hypermethylated (RELN) or histone methylated (SYN1-3, CX30, CX43) in suicide cases compared to healthy controls suggesting that neurotransmission may be compromised in suicide. (3) HPA axis: two genes differentially involved in cortisol release following stress (NR3C1, SKA2) have been found to be both significantly hypermethylated in suicide cases relative to controls supporting the hypothesis of suicide-associated HPA axis dysregulation. (4) Neurotrophic Signalling: several studies examining BDNF and TrkB in post-mortem suicide brains revealed distinct epigenetic regulation pattern for these two genes: the receptor TrkB seems to be regulated via miRNAs (miR-30a, miR-185, miR-195 and miR-49) and histone acetylation while the expression levels of the neurotrophin Bdnf correlate with DNA hypermethylation in the promoter region of the gene.
Table 1 Aberrant epigenetic changes associated with suicidal behavior

Table 2 Differentially expressed miRNAs associated with suicidal behavior
2.1 HPA Stress Axis
It is well established that perturbations in the HPA axis are associated with the pathophysiology of suicide [62]. The HPA axis is a neuroendocrine system, key regulator of cortisol release and the stress response [63]. Research has shown that the HPA axis may be epigenetically programmed by the early-life environment [64]. Several HPA-axis coupled genes have been identified over the years: corticotropin releasing hormone (CRH), corticotropin releasing hormone binding protein (CRHBP), corticotropin releasing hormone receptor 1 (CRHR1), corticotropin releasing hormone receptor 2 (CRHR2), FK506-binding protein 51 (FKBP5) and the glucocorticoid receptor (NR3C1)[65].
More recently, a study aiming to identify DNA methylation shifts linked to severity of the SA in HPA-axis coupled genes was conducted in whole blood derived DNA. The authors reported reduced levels of DNA methylation at two loci within the CRH promoter in serious suicide attempters. Interestingly, significant blood-brain correlations in DNA methylation were also found suggesting that these alterations may impact on expression profile of CRH in the brain [65].
Glucocorticoids. Glucocorticoids play a crucial role in the maintenance and survival of neurons and in synaptic plasticity. A study examining the DNA methylation status of GR exon 1F variant in individuals who died by suicide and were severely abused during childhood observed increased DNA methylation at the GR promoter compared to suicide victims with no childhood abuse or controls [66]. Moreover, GR methylation status appears to regulate the binding of the NGFI-A transcription factor associated with the GR expression [66,67]. The disrupted GR function may result in inadequate control of the HPA axis, possibly leading to hyperactive cortisol secretion and development of anxiety traits. In turn, anxiety mediates the relationship between the exposure to early life adversity (ELA) and SB [64],[68]. More recently, a study has suggested a role for dietary deficiencies in mediating the interaction between an altered GR system and SB. DNA methyltransferases catalyse the transfer of a methyl group from the methyl donor, s-adenoslymethionine (SAM), onto the 5C position of the dinucleotide sequence CG. However, the synthesis of SAM is dependent on the availability of dietary foliates, vitamin B12 and choline, suggesting an alternative mechanism to altering the expression of the GRII exon 17 promoter [69].
SKA2 - Altered microtubule binding. An exciting new candidate in the relationship between cortisol regulation and suicide is the spindle and kinetochore associated protein 2 (SKA2), a gene that has been implicated in GR signalling [70]. This microtubule binding protein is thought to interact with the HPA axis by chaperoning the GR from the cytoplasm to the nucleus upon cortisol binding [70]. Once in the nucleus, the GR can interact with genomic DNA and influence gene expression involved in negative feedback regulation of the HPA axis response. In 2014, a study identified site and allele-specific DNA methylation patterns in the SKA2 gene, in particular increased SKA2 3′ untranslated region methylation and concomitantly decreased SKA2 mRNA levels, were detected in the frontal cortex of suicide completers [71]. Further research found that SKA2 DNA methylation predicted lifetime SA in saliva and blood in individuals suffering from post-traumatic stress disorder (PTSD) [72]. Higher DNA methylation at the same site and allele was also found to predict lower levels of miR-301a in the cortex of depressed suicide completers [73]. Since the expression of this miRNA is tied to SKA2 expression, it was suggested as a proxy of suicide-associated SKA2 decreases [55,72]. Further support to these preliminary findings has been provided by more recent studies suggesting the influence of trauma exposure on SKA2 DNA methylation as well as DNA methylation of the SKA2 gene as a biomarker of suicide risk and stress-related psychopathology [74,75].
2.2 Polyamine System and SAT1
The polyamine system is another stress response pathway which has been extensively characterized in relation to suicide risk. Polyamines, aliphatic compounds with multiple amine groups, have been implicated in regulation of gene expression at transcriptional and posttranscriptional levels, regulating the function of several neuromodulators and acting as neurotransmitters themselves [76,77]. Involvement of epigenetic modifications in the regulation of genes associated with polyamine biosynthesis has been implicated in SB [78].
SAT1 is an enzyme involved in polyamine catabolism and is arguably one of the most consistently downregulated gene in depressed suicide completers [79,80,81,82,83,84,85]. A potential mechanism for SAT1 downregulation is through epigenetic control, as studies have identified that SAT1 promoter DNA methylation is inversely correlated with SAT1 expression. Moreover, there is evidence for histone modifications affecting SAT1 expression along with other key enzymes in polyamine synthesis [54,78,82] and that miRNAs can target polyamine transcripts, including SAT1 [86]. SAT1 has therefore emerged as a potential biomarker for suicide, topping the lists of candidate genes in several studies [84,87,88]. However, it is still unknown whether this gene is involved in major depressive disorder (MDD) independently of suicide, which isoforms are dysregulated and whether the gene undergoes differential splicing in suicide and depression [89].
The expression of several other polyamine-associated enzymes were found altered in the cortex of post-mortem suicide cases. The spermine oxidase (SMOX) gene, encoding for a catabolic enzyme, was found hypermethylated in the promoter region in the prefrontal cortex (PFC) of suicide subjects. However, there was no correlation between aberrant promoter DNA hypermethylation and gene expression changes [83]. Two studies have identified DNA hypomethylation at the promoter region of the arginase 2 (ARG2) and adenosylmethionine decarboxylase1 (AMD1) genes, which correlated with increased gene expression in suicide completers compared to controls [54,78]. A different epigenetic modification showed to regulate the Ornithine Decarboxylase Antizyme 1 (OAZ1) gene, also involved in the intracellular regulation of the polyamine levels. Increased H3K4me3 levels in the promoter region of OAZ1 were found in suicide completers and appeared to correlate with the expression of OAZ1 and ARG2 in Brodmann area 44 [54]. Taken together this research provides support for the involvement of epigenetic modifications in the regulation of genes associated with polyamine biosynthesis, which may play an important role in SB.
2.3 Neurotrophic Pathways - BDNF and NTRK2
Genome-wide DNA methylation studies in the brains of suicide completers indicates that suicide is associated with widespread changes in DNA methylation patterns of neurotrophic and neuroprotective factors in the hippocampus and PFC [40,90]. Brain derived neurotrophic factor (BDNF) is a well-known receptor binding factor with growth factor activity; it is essential for the survival and plasticity of cortical and striatal neurons and it has been implicated in both stress response [17] and mood disorders [91]. Increased BDNF promoter/exon 4 DNA methylation has been reported in suicide brains [92], a finding that is consistent with those observed in depressed patients with a history of SA, or with SI during treatment with antidepressants [93]. Similarly, another study focusing on the Wernicke area of suicide subjects, reported a remarkable increase of DNA methylation at the BDNF promoter IV in suicide subjects with respect to controls [43]. Taken together this evidence suggest that BDNF DNA methylation status maybe a proxy marker for previous suicidal attempts and a clinical biomarker for poor treatment outcomes of SI in depression. However, larger studies are required to confirm these findings.
Interestingly, TrkB, main receptor of BDNF and encoded by the NTRK2 gene, is also regulated through epigenetic changes that appear to have an impact on suicide risk. In brain tissue from individuals who died by suicide, mRNA expression of Trk-BT1, the astrocyte-enriched TrkB truncated variant, was found significantly decreased and correlated with increased DNA methylation at the TrkB-T1 promoter suggesting that astrocytic dysfunction may occur in individuals who die by suicide [53,94,95].
3. Non-Coding RNAs and Their Role in SB
Recently, a number of studies have implicated a role for miRNA dysregulation in the pathogenesis of SB (see Table 2 for full list of suicide-associated miRNAs). Interestingly, some of the suicide-associated genes previously discussed as showing altered DNA methylation and/or histone modifications were found to be targets of differentially expressed miRNAs. For example, the expression levels of two genes involved in the polyamine metabolism (SAT1 and SMOX) were found to be downregulated by overexpression of miRNAs (e.g. miR-139, miR-34c, miR-195, miR-320c) [86,89] in suicide cases compared to controls. Similarly, miR-185, which is thought to regulate the NTRK2 gene (TrkB receptor) is differentially expressed in the frontal cortex of suicide completers [96]. Furthermore, miR-185, together with miR-195 and miR-30a have been implicated in regulating the BDNF signalling pathway [56,96,97], a pathway widely reported as involved in SB. Taken together, these findings suggest a co-ordinated epigenetic and miRNA regulation of pathways relevant to SB.
Sun et al, implicated a role for notch signalling in SI in a study of miRNAs from peripheral blood leukocytes of Chinese MDD patients [98].The expression levels of two notch-associated miRNAs, miR-34b–5p and miR-369–3p, were found to be significantly lower in patients with SI relative to patients without SI [98]. Consistent with these results, expression levels of these miRNAs have been previously shown to be reduced in PFC of depressive suicidal patients compared to normal controls [73], highlighting the potential utility in examining changes in miRNA expression in blood in individuals with SB.
In addition to miRNAs, expression levels of lncRNAs have also been implicated in suicide. Recently, our group implicated a role for differential DNA methylation at the PSORS1C3 locus, a non-coding RNA, in the brains of suicide completers. Although the function of the PSORS1C3 gene product remains unknown, it is thought to potentially regulate nearby genes (for example, POU5F1 and HLA-C), suggesting a role in immune system regulation. Moreover, PSORS1C3 is a known psoriasis susceptibility gene further supporting a role in immune system regulation [34]. Futhermore, a recent study identified six lncRNAs (see Table 2) whose expression was downregulated in peripheral blood samples of MDD patients with SI [99] . Finally, Punzi et al showed that the expression of an uncharacterized lncRNA, LOC285758, is significantly increased in violent suicide cases compared to non-violent suicide cases [100]. Interestingly, DNA methylation of an intragenic CpG island in the myristoylated alanine-rich protein kinase C substrate (MARCKS) gene, a gene previously reported [84] as potential blood biomarker for suicidality, was found to be associated with the expression of this lncRNA, suggesting that the previously reported MARCKS association with violent suicide involves regulation by LOC285758 expression. In conclusion, profiling ncRNAs can substantially contribute to our understanding of how gene expression networks are reorganized in suicide.
4. Current Methodological Caveats in Suicide Epigenetic Research
4.1 Phenotype Assessment
The wide range of phenotypes that may be considered in studies investigating SB further complicate identification of clear biological markers by hampering the comparability of studies, even among studies adopting similar approaches. SI and SA may at times be studied concurrently. These phenotypes are often considered to exist on a spectrum and, as a result, are frequently studied and reported on together in genetic and epigenetic studies. A genetic component has been demonstrated for SA and completed suicide but the role of genetics in SI is currently unclear [11]. Thus, it has been suggested to exclude SI from future SB epigenetic studies [3]. SA ranges from highly lethal, to low lethality for failed SA, and could be divided according to impulsive characteristics or chronic SA. However these two phenotypes, completed suicide and SA, are likely to only partly share underlying etiological and neurobiological mechanisms [101] suggesting therefore the need for a revised and more accurate definition of SA phenotype, which includes lethality, in order to ensure inter-study comparability. Finally, SB is generally a complication of a psychiatric disorders like MDD, schizophrenia and/or bipolar disorder. How to distinguish suicide diathesis-related epigenetic changes from those associated with mood disorders and other suicide-associated psychiatric diseases is a crucial issue. Future epigenetic studies need to examine potential SB-related epigenetic modification in a suicidal population with and without psychiatric disorder comorbidity.
4.2 Tissue Heterogeneity
Compared with genetic studies, epigenetics studies are constrained by several additional biological and methodological design issues. A major challenge that distinguishes epigenetic and genetic studies is the choice of tissue type [25]. In contrast to genetic studies, epigenetic marks are by definition tissue and cell type specific, thus tissue choice for epigenetic studies of complex traits, requires careful consideration [25]. As almost all tissues consist of multiple epigenetically distinct cell types, a major concern in epigenetic epidemiology studies is that any apparent disease-associated epigenetic differences may simply reflect differences in cellular composition [25]. Furthermore, failure to account for this cellular heterogeneity could limit the power of epigenetic studies in complex phenotypes. For whole blood, routine cell counts [102] or the use of algorithms that can infer cellular composition from epigenomic data [103] can be applied to control for this variation statistically, and similar approaches have been developed for other heterogeneous tissues, such as the brain [104]. Approaches such as laser capture microdissection (LCM) or using fluorescence-activated cell sorting (FACS) have been recruited to detect disease-associated changes that are manifest in only a small subset of cells, which may not be detectable in analyses of whole tissue. However, manual sorting [105] and LCM [106] are useful for isolating small numbers of cells but do not provide enough material for epigenomic studies. FACS can isolate larger numbers of cells but may be challenging in tissues such as the adult brain, where cells are morphologically complex and densely interconnected. To overcome these challenges nuclei can be isolated from post-mortem tissue and then sorted using FACS for cell type specific epigenomic /transcriptomic studies [107,108,109,110]. However, FACS-sorted nuclei are fragile and difficult to concentrate into the small volumes that are optimal for chromatin assays or to keep intact for DNA methylation assays. Another major challenge is selecting nuclear expressed cell-type specific markers for sorting. Although several cell-specific markers have been identified and validated over the years, most of them are expressed in the cytoplasm or on the cellular membrane making them unsuitable for nuclei sorting. Neuronal nuclear antigen (NeuN) is commonly used as a robust marker of neurons and its application in flow cytometry analyses have been widely validated by several recent studies in mouse and human post-mortem brain samples [104,111,112,113,114]. Although NeuN is specific to neurons, not all neurons express NeuN (e.g. cerebellar Purkinje neurons, olfactory bulb mitral cells, cortical Cajal-Retzius neurons, neurons of the inferior olive and dentate nuclei) [115,116]. Moreover, no robust nuclear markers suitable for FACS are available to date for mature oligodendrocytes, astrocytes and microglia isolation from human brain tissue.
Further research aiming to identify new markers for nuclei labelling is needed to separate nuclei from different brain regions as well as different cell-types and neuronal sub-types unique to different cortical layers. This would empower research discoveries on nuclear changes occurring during disease progression, especially in rare but important populations, such as immune cell types present in the brain, which are thought to be critical in neuroprotective and neuroinflammatory processes [117].
4.3 True Methylation Assessment
5hmC plays a key role in the brain, where it is particularly abundant and dynamic during development [118,119]. Current molecular approaches used to measure levels of DNA methylation (including sodium bisulphite conversion methods) do not distinguish between 5mC and 5hmC [120] suggesting that densely hydroxymethylated regions of DNA may be underrepresented in quantitative methylation analyses. However, several methodologies have now been developed to address this issue (including oxBS-Seq [121], mTAB-Seq [122] or Aba-Seq [123]). 5mC and 5hmC seem to have opposite relationships with transcriptional activity, with 5mC negatively correlating with gene expression [124] and 5hmC positively correlating with gene expression in the rodent [125] and human brains [118]. Although many studies showed that 5hmC is linked with neurological disorders such as Alzheimer’s and Huntington’s diseases [126,127] and psychiatric disorders disorder [128,129,130] its potential implication in SB remains unknown.
4.4 Caveats of miRNA Profiling
Expression profiling of miRNAs have increased our understanding of which miRNAs are present in specific tissues and how they may change under pathological conditions [131]. However, once identified, linking a miRNA to its mRNA targets can be a challenging task, and the mRNA target pairs should be validated. As reviewed elsewhere [132], a very small fraction of software-predicted miRNA targets are validated in vivo and many databases for miRNA target prediction are being developed in order to address this issue [132]. However, those databases can provide a surprising level of divergent results when we take into account degrees of sequence similarity, conservation, site accessibility, and variation in the targeted regions of the mRNA. Similarly, there are a number of conflicting studies with regard to the magnitude and direction of biologically-relevant miRNA expression changes in psychiatric disorders [133,134]. This could be due to tissue-specific variation in expression levels as well as heterogeneity in quantification and normalization procedures [135]. Furthermore, some studies on miRNAs and depression were conducted in peripheral blood despite uncertainties regarding how closely changes in peripheral miRNA expression reflect modifications in the central nervous system (e.g., [136]). Finally, it should be also considered that “control” RNAs commonly used to normalize miRNA data (U6, U44, and U48) are very sensitive to post-mortem decay [132].
4.5 Sample Size and Peripheral Tissue Profiling
The abundance and availability of brain samples is a big challenge and many studies are relying on a limited number of brain samples reducing the power of epigenome-wide association studies [40]. Only genes that have been replicated in independent methylation studies should be considered further as primary candidate genes. Although it is desirable to increase sample sizes in the next few years, it is unfeasible to reach the numbers of samples currently being analysed in GWASs using human brain [40]. The small sample size of most of the current studies also represents a limitation when it comes to identifying gender-specific epigenetic changes associated to SB. Studies are looking towards the use of peripheral tissues (e.g. whole blood, saliva, cerebrospinal fluid (CSF)) to model a brain-related phenotype. Given the tissue-specific nature of the epigenome, the assessment of disease-relevant tissue is an important consideration for EWASs. Although some studies suggest that peripheral samples (e.g. CSF) would adequately model brain gene expression changes, little is known about whether easily accessible tissues, such as whole blood, can be used to address questions about inter-individual epigenomic variation in inaccessible tissues, such as the brain [137]. Epigenetic marks are more variable between different tissues of the same individual than between the same tissue of different individuals. There is however evidence of within-individual epigenetic variation correlation across tissues [138]. A recent study explored co-variation between tissues and the extent to which methylomic variation in blood is predictive of inter-individual variation identified in the brain. Their data suggest that for the majority of the genome, a blood-based EWAS for disorders where brain is presumed to be the primary tissue of interest will give limited information relating to underlying pathological processes [137]. However, the authors do not discount the utility of using a blood-based EWASs to identify biomarkers of disease phenotypes manifest in the brain [137].
4.6 Animal Models
To date no convincing animal models of suicide have been produced [4]. Suicidal behaviour is challenging to detail in a non-human model as the underlying pathophysiology is poorly known and the risk for suicidal acts is multi‐factorial, consisting of a range of biological (e.g. genetics), psychiatric, psychosocial, interpersonal and cultural risk factors. The most promising endophenotypes worth investigating in animals are the cortisol social-stress response and the aggression/impulsivity trait, involving the serotonergic system as well as neurotrophic factors and neurotrophins which have been implicated in suicide previously [4]. Future animal studies could focus on elucidating the effect of suicide-associated epigenetic changes on gene function and how this might impact known suicide-associated endophenotypes and biological pathways.
4.7 Causality
Currently it remains unclear whether the epigenetic changes observed cause SB and suicidal acts or if these phenotypes (or underlying psychiatric conditions/medication) cause the detected epigenetic changes. In the attempt to answer this question, Mendelian randomization (MR) is proving to be a promising approach. MR posits that if a biomarker is causally related to a disease or phenotype, genetic variant(s) controlling activity of that biomarker should also be associated with the outcome [139,140]. Using genetic variants as proxy for exposure overcomes confounding because genetic variants are inherited at random during meiosis, so they are unrelated to potential confounders (measured or unmeasured). Therefore, following the principles of MR may represent a valid method for revealing the role of specific genes as modifiers/risk factors of SB, leading to a possible association between suicidality and a given biomarker. Future studies examining epigenetic alterations in SB should be undertaken by applying these novel approaches in order to investigate the direction of effect. For instance, the 2-step epigenetic MR approach, which is an extension of MR assumptions, allows researchers to investigate the causal role of DNA methylation in the association between an environmental exposure and SB [141,142].
None of the studies outlined in this review have attempted to examine the causal pathways between epigenetics changes and SB, either statistically or experimentally. Recent advances in CRISPR-Cas9 based systems enable researchers to direct epigenetic machinery to specific locations with possibly greater ease and at a lower cost. Once the major drawbacks with dCas9 have been solved (such as the inefficiency of precise base editing and off-target activities), in vivo epigenome editing methods will potentially allow for more labs to investigate the behavioural outcomes of specific epigenetic patterns in cell or animal models. Although most of the excitement around CRISPR to date has been focused on the ability of genome editing to cure disease, epigenome editing methods might enable more subtle and reversible modulatory control [24]. Future studies will greatly benefit from these new tools and allow for a precise investigation of the direct molecular and behavioural outcomes of suicide -induced epigenetic patterns in the brain.
4.8 Other Considerations
Typically, post-mortem studies report post-mortem interval (PMI), which represents the amount of time between a subject’s death and collection and processing of the brain. It has previously been shown that DNA, miRNAs and some histone modifications are stable across extended PMIs [143,144,145,146]. Moreover, a recent study has reported that neither global nor site-specific levels of 5mC and 5hmC are affected by the post-mortem intervals [46].
A further complication stems from the emergence of SI during antidepressant treatment. There is indeed a longstanding belief that antidepressants might have an early “triggering effect” that induce depressed patients to pursue suicidal impulses before the treatment-induced mood improvement [147]. Some patients being treated for depression and other psychiatric illnesses experience suicidal thoughts and actions (suicidality) as previously reported in a small proportion of patients receiving selective serotonin reuptake inhibitors [147]. Therefore, when medication data are not available for all individuals included in a study, the possibility that the observed epigenetic changes are confounded by medication cannot be ruled out.
5. Future Perspectives
A multi-omics approach to disentangle the molecular basis underlying SB is warranted to further our understanding of suicidality and SB. Genome-wide investigations have demonstrated that the genetic predisposition to complex traits is highly polygenic. The common variants detected by polygenic risk score analyses may help to predict the extent to which the genetic predisposition for suicidal acts has influence on the risk of suicide attempts. By integrating polygenic risk scores for SA, DNA modifications, chromatin modifications and miRNA expression profiling together would allow the research community to gain a broader and deeper understanding of the pathways primarily involved in the neurobiology of suicide and their potential co-ordinated interaction. For the same reason, future studies should also examine the transcriptional consequences of the observed suicide-related DNA methylation changes. System biology methods, such as weighted correlation network analysis (WGCNA), could be applied to identify suicide-associated gene pathways and networks that could be further targeted as biomarkers or therapeutic targets for suicide prevention.
Author Contributions
All authors conceived the topic of the review. SP prepared the first draft of the review and ELD and TMM edited subsequent versions of the draft. SP and TMM prepared and conceived Figure 1 and Figure 2.
Competing Interests
The authors have declared that no competing interests exist.
References
- Report CH. CDC 2017 [Available from: https://www.cdc.gov/.
- Organization WH. 2017 [Available from: http://www.who.int/news-room/fact-sheets/detail/suicide.
- Tsai SJ, Hong CJ, Liou YJ. Recent molecular genetic studies and methodological issues in suicide research. Prog Neuropsychopharmacol Biol Psychiatry. 2011; 35: 809-817. [CrossRef] [Google scholar] [PubMed]
- Preti A. Animal model and neurobiology of suicide. Prog Neuropsychopharmacol Biol Psychiatry. 2011; 35: 818-830. [CrossRef] [Google scholar] [PubMed]
- Nock MK, Green JG, Hwang I, McLaughlin KA, Sampson NA, Zaslavsky AM, et al. Prevalence, correlates, and treatment of lifetime suicidal behavior among adolescents: results from the National Comorbidity Survey Replication Adolescent Supplement. JAMA Psychiatry. 2013; 70: 300-310. [CrossRef] [Google scholar] [PubMed]
- Roy A, Segal NL. Suicidal behavior in twins: a replication. J Affect Disord. 2001; 66: 71-74. [CrossRef] [Google scholar] [PubMed]
- Mullins N, Perroud N, Uher R, Butler AW, Cohen-Woods S, Rivera M, et al. Genetic relationships between suicide attempts, suicidal ideation and major psychiatric disorders: a genome-wide association and polygenic scoring study. Am J Med Genet B Neuropsychiatr Genet. 2014; 165B: 428-437. [CrossRef] [Google scholar] [PubMed]
- Fu Q, Heath AC, Bucholz KK, Nelson EC, Glowinski AL, Goldberg J, et al. A twin study of genetic and environmental influences on suicidality in men. Psychol Med. 2002; 32: 11-24. [CrossRef] [Google scholar] [PubMed]
- Statham DJ, Heath AC, Madden PA, Bucholz KK, Bierut L, Dinwiddie SH, et al. Suicidal behaviour: an epidemiological and genetic study. Psychol Med. 1998; 28: 839-855. [CrossRef] [Google scholar] [PubMed]
- Pedersen NL, Fiske A. Genetic influences on suicide and nonfatal suicidal behavior: twin study findings. Eur Psychiatry. 2010; 25: 264-267. [CrossRef] [Google scholar] [PubMed]
- Brent DA, Bridge J, Johnson BA, Connolly J. Suicidal behavior runs in families. A controlled family study of adolescent suicide victims. Arch Gen Psychiatry. 1996; 53: 1145-1152. [CrossRef] [Google scholar] [PubMed]
- Brent DA, Oquendo M, Birmaher B, Greenhill L, Kolko D, Stanley B, et al. Familial transmission of mood disorders: convergence and divergence with transmission of suicidal behavior. J Am Acad Child Adolesc Psychiatry. 2004; 43: 1259-1266. [CrossRef] [Google scholar] [PubMed]
- Kim CD, Seguin M, Therrien N, Riopel G, Chawky N, Lesage AD, et al. Familial aggregation of suicidal behavior: a family study of male suicide completers from the general population. Am J Psychiatry. 2005; 162: 1017-1019. [CrossRef] [Google scholar] [PubMed]
- Anguelova M, Benkelfat C, Turecki G. A systematic review of association studies investigating genes coding for serotonin receptors and the serotonin transporter: II. Suicidal behavior. Mol Psychiatry. 2003; 8: 646-653. [CrossRef] [Google scholar] [PubMed]
- Brezo J, Klempan T, Turecki G. The genetics of suicide: a critical review of molecular studies. Psychiatr Clin North Am. 2008; 31: 179-203. [CrossRef] [Google scholar] [PubMed]
- Mann JJ. The serotonergic system in mood disorders and suicidal behaviour. Philos Trans R Soc Lond B Biol Sci. 2013; 368: 20120537. [CrossRef] [Google scholar] [PubMed]
- Dwivedi Y. Brain-derived neurotrophic factor and suicide pathogenesis. Ann Med. 2010; 42: 87-96. [CrossRef] [Google scholar] [PubMed]
- Schosser A, Butler AW, Ising M, Perroud N, Uher R, Ng MY, et al. Genomewide association scan of suicidal thoughts and behaviour in major depression. PLoS One. 2011; 6: e20690. [CrossRef] [Google scholar] [PubMed]
- Willour VL, Seifuddin F, Mahon PB, Jancic D, Pirooznia M, Steele J, et al. A genome-wide association study of attempted suicide. Mol Psychiatry. 2012; 17: 433-444. [CrossRef] [Google scholar] [PubMed]
- Galfalvy H, Zalsman G, Huang YY, Murphy L, Rosoklija G, Dwork AJ, et al. A pilot genome wide association and gene expression array study of suicide with and without major depression. World J Biol Psychiatry. 2013; 14: 574-582. [CrossRef] [Google scholar] [PubMed]
- Turecki G, Brent DA. Suicide and suicidal behaviour. Lancet. 2016; 387: 1227-1239. [CrossRef] [Google scholar] [PubMed]
- Bell JT, Tsai PC, Yang TP, Pidsley R, Nisbet J, Glass D, et al. Epigenome-wide scans identify differentially methylated regions for age and age-related phenotypes in a healthy ageing population. PLoS Genet. 2012; 8: e1002629. [CrossRef] [Google scholar] [PubMed]
- Feil R, Fraga MF. Epigenetics and the environment: emerging patterns and implications. Nat Rev Genet. 2012; 13: 97-109. [CrossRef] [Google scholar] [PubMed]
- Burns SB, Szyszkowicz JK, Luheshi GN, Lutz PE, Turecki G. Plasticity of the epigenome during early-life stress. Semin Cell Dev Biol. 2018; 77: 115-132. [CrossRef] [Google scholar] [PubMed]
- Mill J, Heijmans BT. From promises to practical strategies in epigenetic epidemiology. Nat Rev Genet. 2013; 14: 585-594. [CrossRef] [Google scholar] [PubMed]
- Wong CC, Meaburn EL, Ronald A, Price TS, Jeffries AR, Schalkwyk LC, et al. Methylomic analysis of monozygotic twins discordant for autism spectrum disorder and related behavioural traits. Mol Psychiatry. 2014; 19: 495-503. [CrossRef] [Google scholar] [PubMed]
- Pidsley R, Viana J, Hannon E, Spiers H, Troakes C, Al-Saraj S, et al. Methylomic profiling of human brain tissue supports a neurodevelopmental origin for schizophrenia. Genome Biol. 2014; 15: 483. [CrossRef] [Google scholar] [PubMed]
- Uddin M, Aiello AE, Wildman DE, Koenen KC, Pawelec G, de Los Santos R, et al. Epigenetic and immune function profiles associated with posttraumatic stress disorder. Proc Natl Acad Sci U S A. 2010; 107: 9470-9475. [CrossRef] [Google scholar] [PubMed]
- Fisher HL, Murphy TM, Arseneault L, Caspi A, Moffitt TE, Viana J, et al. Methylomic analysis of monozygotic twins discordant for childhood psychotic symptoms. Epigenetics. 2015; 10: 1014-1023. [CrossRef] [Google scholar] [PubMed]
- Murphy TM, O'Donovan A, Mullins N, O'Farrelly C, McCann A, Malone K. Anxiety is associated with higher levels of global DNA methylation and altered expression of epigenetic and interleukin-6 genes. Psychiatr Genet. 2015; 25: 71-78. [CrossRef] [Google scholar] [PubMed]
- Dempster EL, Wong CC, Lester KJ, Burrage J, Gregory AM, Mill J, et al. Genome-wide methylomic analysis of monozygotic twins discordant for adolescent depression. Biol Psychiatry. 2014; 76: 977-983. [CrossRef] [Google scholar] [PubMed]
- Davies MN, Krause L, Bell JT, Gao F, Ward KJ, Wu H, et al. Hypermethylation in the ZBTB20 gene is associated with major depressive disorder. Genome Biol. 2014; 15: R56. [CrossRef] [Google scholar] [PubMed]
- Uddin M, Koenen KC, Aiello AE, Wildman DE, de los Santos R, Galea S. Epigenetic and inflammatory marker profiles associated with depression in a community-based epidemiologic sample. Psychol Med. 2011; 41: 997-1007. [CrossRef] [Google scholar] [PubMed]
- Murphy TM, Crawford B, Dempster EL, Hannon E, Burrage J, Turecki G, et al. Methylomic profiling of cortex samples from completed suicide cases implicates a role for PSORS1C3 in major depression and suicide. Transl Psychiatry. 2017; 7: e989. [CrossRef] [Google scholar] [PubMed]
- Murphy TM, Mullins N, Ryan M, Foster T, Kelly C, McClelland R, et al. Genetic variation in DNMT3B and increased global DNA methylation is associated with suicide attempts in psychiatric patients. Genes Brain Behav. 2013; 12: 125-132. [CrossRef] [Google scholar] [PubMed]
- Goldberg AD, Allis CD, Bernstein E. Epigenetics: a landscape takes shape. Cell. 2007; 128: 635-638. [CrossRef] [Google scholar] [PubMed]
- Labonte B, Turecki G. The epigenetics of suicide: explaining the biological effects of early life environmental adversity. Arch Suicide Res. 2010; 14: 291-310. [CrossRef] [Google scholar] [PubMed]
- Autry AE, Monteggia LM. Epigenetics in suicide and depression. Biol Psychiatry. 2009; 66: 812-813. [CrossRef] [Google scholar] [PubMed]
- McGowan PO, Szyf M. The epigenetics of social adversity in early life: implications for mental health outcomes. Neurobiol Dis. 2010; 39: 66-72. [CrossRef] [Google scholar] [PubMed]
- Schneider E, El Hajj N, Muller F, Navarro B, Haaf T. Epigenetic Dysregulation in the Prefrontal Cortex of Suicide Completers. Cytogenet Genome Res. 2015; 146: 19-27. [CrossRef] [Google scholar] [PubMed]
- He Y, Ecker JR. Non-CG Methylation in the Human Genome. Annu Rev Genomics Hum Genet. 2015; 16: 55-77. [CrossRef] [Google scholar] [PubMed]
- Lister R, Mukamel EA, Nery JR, Urich M, Puddifoot CA, Johnson ND, et al. Global epigenomic reconfiguration during mammalian brain development. Science. 2013; 341: 1237905. [CrossRef] [Google scholar] [PubMed]
- Keller S, Sarchiapone M, Zarrilli F, Tomaiuolo R, Carli V, Angrisano T, et al. TrkB gene expression and DNA methylation state in Wernicke area does not associate with suicidal behavior. J Affect Disord. 2011; 135: 400-404. [CrossRef] [Google scholar] [PubMed]
- Haghighi F, Xin Y, Chanrion B, O'Donnell AH, Ge Y, Dwork AJ, et al. Increased DNA methylation in the suicide brain. Dialogues Clin Neurosci. 2014; 16: 430-438. [Google scholar]
- Tucker KL. Methylated cytosine and the brain: a new base for neuroscience. Neuron. 2001; 30: 649-652. [CrossRef] [Google scholar] [PubMed]
- Gross JA, Nagy C, Lin L, Bonneil E, Maheu M, Thibault P, et al. Global and Site-Specific Changes in 5-Methylcytosine and 5-Hydroxymethylcytosine after Extended Post-mortem Interval. Front Genet. 2016; 7: 120. [CrossRef] [Google scholar] [PubMed]
- Berger SL. The complex language of chromatin regulation during transcription. Nature. 2007; 447: 407-412. [CrossRef] [Google scholar] [PubMed]
- Kriaucionis S, Heintz N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science. 2009; 324: 929-930. [CrossRef] [Google scholar] [PubMed]
- Tahiliani M, Koh KP, Shen Y, Pastor WA, Bandukwala H, Brudno Y, et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science. 2009; 324: 930-935. [CrossRef] [Google scholar] [PubMed]
- Ito S, Shen L, Dai Q, Wu SC, Collins LB, Swenberg JA, et al. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science. 2011; 333: 1300-1303. [CrossRef] [Google scholar] [PubMed]
- Szulwach KE, Li X, Li Y, Song CX, Wu H, Dai Q, et al. 5-hmC-mediated epigenetic dynamics during postnatal neurodevelopment and aging. Nat Neurosci. 2011; 14: 1607-1616. [CrossRef] [Google scholar] [PubMed]
- Hahn MA, Qiu R, Wu X, Li AX, Zhang H, Wang J, et al. Dynamics of 5-hydroxymethylcytosine and chromatin marks in Mammalian neurogenesis. Cell Rep. 2013; 3: 291-300. [CrossRef] [Google scholar] [PubMed]
- Ernst C, Chen ES, Turecki G. Histone methylation and decreased expression of TrkB.T1 in orbital frontal cortex of suicide completers. Mol Psychiatry. 2009; 14: 830-832. [CrossRef] [Google scholar] [PubMed]
- Fiori LM, Gross JA, Turecki G. Effects of histone modifications on increased expression of polyamine biosynthetic genes in suicide. Int J Neuropsychopharmacol. 2012; 15: 1161-1166. [CrossRef] [Google scholar] [PubMed]
- Smalheiser NR, Lugli G, Zhang H, Rizavi H, Cook EH, Dwivedi Y. Expression of microRNAs and other small RNAs in prefrontal cortex in schizophrenia, bipolar disorder and depressed subjects. PLoS One. 2014; 9: e86469. [CrossRef] [Google scholar] [PubMed]
- Serafini G, Pompili M, Hansen KF, Obrietan K, Dwivedi Y, Shomron N, et al. The involvement of microRNAs in major depression, suicidal behavior, and related disorders: a focus on miR-185 and miR-491-3p. Cell Mol Neurobiol. 2014; 34: 17-30. [CrossRef] [Google scholar] [PubMed]
- Xu B, Karayiorgou M, Gogos JA. MicroRNAs in psychiatric and neurodevelopmental disorders. Brain Res. 2010; 1338: 78-88. [CrossRef] [Google scholar] [PubMed]
- Miller BH, Wahlestedt C. MicroRNA dysregulation in psychiatric disease. Brain Res. 2010; 1338: 89-99. [CrossRef] [Google scholar] [PubMed]
- Derrien T, Johnson R, Bussotti G, Tanzer A, Djebali S, Tilgner H, et al. The GENCODE v7 catalog of human long noncoding RNAs: analysis of their gene structure, evolution, and expression. Genome Res. 2012; 22: 1775-1789. [CrossRef] [Google scholar] [PubMed]
- Aprea J, Prenninger S, Dori M, Ghosh T, Monasor LS, Wessendorf E, et al. Transcriptome sequencing during mouse brain development identifies long non-coding RNAs functionally involved in neurogenic commitment. EMBO J. 2013; 32: 3145-3160. [CrossRef] [Google scholar] [PubMed]
- Spadaro PA, Flavell CR, Widagdo J, Ratnu VS, Troup M, Ragan C, et al. Long Noncoding RNA-Directed Epigenetic Regulation of Gene Expression Is Associated With Anxiety-like Behavior in Mice. Biol Psychiatry. 2015; 78: 848-859. [CrossRef] [Google scholar] [PubMed]
- Mann JJ, Currier DM. Stress, genetics and epigenetic effects on the neurobiology of suicidal behavior and depression. Eur Psychiatry. 2010; 25: 268-271. [CrossRef] [Google scholar] [PubMed]
- Raison CL, Miller AH. When not enough is too much: the role of insufficient glucocorticoid signaling in the pathophysiology of stress-related disorders. Am J Psychiatry. 2003; 160: 1554-1565. [CrossRef] [Google scholar] [PubMed]
- Turecki G. The molecular bases of the suicidal brain. Nat Rev Neurosci. 2014; 15: 802-816. [CrossRef] [Google scholar] [PubMed]
- Jokinen J, Bostrom AE, Dadfar A, Ciuculete DM, Chatzittofis A, Asberg M, et al. Epigenetic Changes in the CRH Gene are Related to Severity of Suicide Attempt and a General Psychiatric Risk Score in Adolescents. EBioMedicine. 2018; 27: 123-133. [CrossRef] [Google scholar] [PubMed]
- McGowan PO, Sasaki A, D'Alessio AC, Dymov S, Labonte B, Szyf M, et al. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat Neurosci. 2009; 12: 342-348. [CrossRef] [Google scholar] [PubMed]
- Weaver IC, Cervoni N, Champagne FA, D'Alessio AC, Sharma S, Seckl JR, et al. Epigenetic programming by maternal behavior. Nat Neurosci. 2004; 7: 847-854. [CrossRef] [Google scholar] [PubMed]
- Wanner B, Vitaro F, Tremblay RE, Turecki G. Childhood trajectories of anxiousness and disruptiveness explain the association between early-life adversity and attempted suicide. Psychol Med. 2012; 42: 2373-2382. [CrossRef] [Google scholar] [PubMed]
- Bennett MR. The prefrontal-limbic network in depression: Modulation by hypothalamus, basal ganglia and midbrain. Prog Neurobiol. 2011; 93: 468-487. [CrossRef] [Google scholar] [PubMed]
- Rice L, Waters CE, Eccles J, Garside H, Sommer P, Kay P, et al. Identification and functional analysis of SKA2 interaction with the glucocorticoid receptor. J Endocrinol. 2008; 198: 499-509. [CrossRef] [Google scholar] [PubMed]
- Guintivano J, Brown T, Newcomer A, Jones M, Cox O, Maher BS, et al. Identification and replication of a combined epigenetic and genetic biomarker predicting suicide and suicidal behaviors. Am J Psychiatry. 2014; 171: 1287-1296. [CrossRef] [Google scholar] [PubMed]
- Kaminsky Z, Wilcox HC, Eaton WW, Van Eck K, Kilaru V, Jovanovic T, et al. Epigenetic and genetic variation at SKA2 predict suicidal behavior and post-traumatic stress disorder. Transl Psychiatry. 2015 ;5: e627. [CrossRef] [Google scholar] [PubMed]
- Smalheiser NR, Lugli G, Rizavi HS, Torvik VI, Turecki G, Dwivedi Y. MicroRNA expression is down-regulated and reorganized in prefrontal cortex of depressed suicide subjects. PLoS One. 2012; 7: e33201. [CrossRef] [Google scholar] [PubMed]
- Boks MP, Rutten BP, Geuze E, Houtepen LC, Vermetten E, Kaminsky Z, et al. SKA2 Methylation is Involved in Cortisol Stress Reactivity and Predicts the Development of Post-Traumatic Stress Disorder (PTSD) After Military Deployment. Neuropsychopharmacology. 2016; 41: 1350-1356. [CrossRef] [Google scholar] [PubMed]
- Sadeh N, Wolf EJ, Logue MW, Hayes JP, Stone A, Griffin LM, et al. Epigenetic Variation at Ska2 Predicts Suicide Phenotypes and Internalizing Psychopathology. Depress Anxiety. 2016; 33: 308-315. [CrossRef] [Google scholar] [PubMed]
- Limon A, Mamdani F, Hjelm BE, Vawter MP, Sequeira A. Targets of polyamine dysregulation in major depression and suicide: Activity-dependent feedback, excitability, and neurotransmission. Neurosci Biobehav Rev. 2016; 66: 80-91. [CrossRef] [Google scholar] [PubMed]
- Gilad GM, Gilad VH. Overview of the brain polyamine-stress-response: regulation, development, and modulation by lithium and role in cell survival. Cell Mol Neurobiol. 2003; 23: 637-649. [CrossRef] [Google scholar] [PubMed]
- Gross JA, Fiori LM, Labonte B, Lopez JP, Turecki G. Effects of promoter methylation on increased expression of polyamine biosynthetic genes in suicide. J Psychiatr Res. 2013; 47: 513-519. [CrossRef] [Google scholar] [PubMed]
- Sequeira A, Gwadry FG, Ffrench-Mullen JM, Canetti L, Gingras Y, Casero RA, Jr., et al. Implication of SSAT by gene expression and genetic variation in suicide and major depression. Arch Gen Psychiatry. 2006; 63: 35-48. [CrossRef] [Google scholar] [PubMed]
- Fiori LM, Mechawar N, Turecki G. Identification and characterization of spermidine/spermine N1-acetyltransferase promoter variants in suicide completers. Biol Psychiatry. 2009; 66: 460-467. [CrossRef] [Google scholar] [PubMed]
- Guipponi M, Deutsch S, Kohler K, Perroud N, Le Gal F, Vessaz M, et al. Genetic and epigenetic analysis of SSAT gene dysregulation in suicidal behavior. Am J Med Genet B Neuropsychiatr Genet. 2009; 150B: 799-807. [CrossRef] [Google scholar] [PubMed]
- Fiori LM, Turecki G. Epigenetic regulation of spermidine/spermine N1-acetyltransferase (SAT1) in suicide. J Psychiatr Res. 2011; 45: 1229-1235. [CrossRef] [Google scholar] [PubMed]
- Fiori LM, Turecki G. Genetic and epigenetic influences on expression of spermine synthase and spermine oxidase in suicide completers. Int J Neuropsychopharmacol. 2010; 13: 725-736. [CrossRef] [Google scholar] [PubMed]
- Le-Niculescu H, Levey DF, Ayalew M, Palmer L, Gavrin LM, Jain N, et al. Discovery and validation of blood biomarkers for suicidality. Mol Psychiatry. 2013; 18: 1249-1264. [CrossRef] [Google scholar] [PubMed]
- Sequeira A, Klempan T, Canetti L, ffrench-Mullen J, Benkelfat C, Rouleau GA, et al. Patterns of gene expression in the limbic system of suicides with and without major depression. Mol Psychiatry. 2007; 12: 640-655. [CrossRef] [Google scholar] [PubMed]
- Lopez JP, Fiori LM, Gross JA, Labonte B, Yerko V, Mechawar N, et al. Regulatory role of miRNAs in polyamine gene expression in the prefrontal cortex of depressed suicide completers. Int J Neuropsychopharmacol. 2014; 17: 23-32. [CrossRef] [Google scholar] [PubMed]
- Niculescu AB, Levey D, Le-Niculescu H, Niculescu E, Kurian SM, Salomon D. Psychiatric blood biomarkers: avoiding jumping to premature negative or positive conclusions. Mol Psychiatry. 2015; 20: 286-288. [CrossRef] [Google scholar] [PubMed]
- Niculescu AB, Levey DF, Phalen PL, Le-Niculescu H, Dainton HD, Jain N, et al. Understanding and predicting suicidality using a combined genomic and clinical risk assessment approach. Mol Psychiatry. 2015; 20: 1266-1285. [CrossRef] [Google scholar] [PubMed]
- Pantazatos SP, Andrews SJ, Dunning-Broadbent J, Pang J, Huang YY, Arango V, et al. Isoform-level brain expression profiling of the spermidine/spermine N1-Acetyltransferase1 (SAT1) gene in major depression and suicide. Neurobiol Dis. 2015; 79: 123-134. [CrossRef] [Google scholar] [PubMed]
- Labonte B, Suderman M, Maussion G, Lopez JP, Navarro-Sanchez L, Yerko V, et al. Genome-wide methylation changes in the brains of suicide completers. Am J Psychiatry. 2013; 170: 511-520. [CrossRef] [Google scholar] [PubMed]
- Phillips C. Brain-Derived Neurotrophic Factor, Depression, and Physical Activity: Making the Neuroplastic Connection. Neural Plast. 2017; 2017: 7260130. [CrossRef] [Google scholar] [PubMed]
- Keller S, Sarchiapone M, Zarrilli F, Videtic A, Ferraro A, Carli V, et al. Increased BDNF promoter methylation in the Wernicke area of suicide subjects. Arch Gen Psychiatry. 2010; 67: 258-267. [CrossRef] [Google scholar] [PubMed]
- Kang HJ, Kim JM, Lee JY, Kim SY, Bae KY, Kim SW, et al. BDNF promoter methylation and suicidal behavior in depressive patients. J Affect Disord. 2013; 151: 679-685. [CrossRef] [Google scholar] [PubMed]
- Ernst C, Deleva V, Deng X, Sequeira A, Pomarenski A, Klempan T, et al. Alternative splicing, methylation state, and expression profile of tropomyosin-related kinase B in the frontal cortex of suicide completers. Arch Gen Psychiatry. 2009; 66: 22-32. [CrossRef] [Google scholar] [PubMed]
- Maussion G, Yang J, Suderman M, Diallo A, Nagy C, Arnovitz M, et al. Functional DNA methylation in a transcript specific 3'UTR region of TrkB associates with suicide. Epigenetics. 2014; 9: 1061-1070. [CrossRef] [Google scholar] [PubMed]
- Maussion G, Yang J, Yerko V, Barker P, Mechawar N, Ernst C, et al. Regulation of a truncated form of tropomyosin-related kinase B (TrkB) by Hsa-miR-185* in frontal cortex of suicide completers. PLoS One. 2012; 7: e39301. [CrossRef] [Google scholar] [PubMed]
- Mellios N, Huang HS, Grigorenko A, Rogaev E, Akbarian S. A set of differentially expressed miRNAs, including miR-30a-5p, act as post-transcriptional inhibitors of BDNF in prefrontal cortex. Hum Mol Genet. 2008; 17: 3030-3042. [CrossRef] [Google scholar] [PubMed]
- Sun N, Lei L, Wang Y, Yang C, Liu Z, Li X, et al. Preliminary comparison of plasma notch-associated microRNA-34b and -34c levels in drug naive, first episode depressed patients and healthy controls. J Affect Disord. 2016; 194: 109-114. [CrossRef] [Google scholar] [PubMed]
- Cui X, Niu W, Kong L, He M, Jiang K, Chen S, et al. Long noncoding RNA expression in peripheral blood mononuclear cells and suicide risk in Chinese patients with major depressive disorder. Brain Behav. 2017; 7: e00711. [CrossRef] [Google scholar] [PubMed]
- Punzi G, Ursini G, Shin JH, Kleinman JE, Hyde TM, Weinberger DR. Increased expression of MARCKS in post-mortem brain of violent suicide completers is related to transcription of a long, noncoding, antisense RNA. Mol Psychiatry. 2014; 19: 1057-1059. [CrossRef] [Google scholar] [PubMed]
- Turecki G, Ernst C, Jollant F, Labonte B, Mechawar N. The neurodevelopmental origins of suicidal behavior. Trends Neurosci. 2012; 35: 14-23. [CrossRef] [Google scholar] [PubMed]
- Heijmans BT, Mill J. Commentary: The seven plagues of epigenetic epidemiology. Int J Epidemiol. 2012; 41: 74-78. [CrossRef] [Google scholar] [PubMed]
- Houseman EA, Accomando WP, Koestler DC, Christensen BC, Marsit CJ, Nelson HH, et al. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinformatics. 2012; 13: 86. [CrossRef] [Google scholar] [PubMed]
- Guintivano J, Aryee MJ, Kaminsky ZA. A cell epigenotype specific model for the correction of brain cellular heterogeneity bias and its application to age, brain region and major depression. Epigenetics. 2013; 8: 290-302. [CrossRef] [Google scholar] [PubMed]
- Sugino K, Hempel CM, Miller MN, Hattox AM, Shapiro P, Wu C, et al. Molecular taxonomy of major neuronal classes in the adult mouse forebrain. Nat Neurosci. 2006; 9: 99-107. [CrossRef] [Google scholar] [PubMed]
- Emmert-Buck MR, Bonner RF, Smith PD, Chuaqui RF, Zhuang Z, Goldstein SR, et al. Laser capture microdissection. Science. 1996; 274: 998-1001. [CrossRef] [Google scholar] [PubMed]
- Kepecs A, Fishell G. Interneuron cell types are fit to function. Nature. 2014; 505: 318-326. [CrossRef] [Google scholar] [PubMed]
- Amin NM, Greco TM, Kuchenbrod LM, Rigney MM, Chung MI, Wallingford JB, et al. Proteomic profiling of cardiac tissue by isolation of nuclei tagged in specific cell types (INTACT). Development. 2014; 141: 962-973. [CrossRef] [Google scholar] [PubMed]
- Henry GL, Davis FP, Picard S, Eddy SR. Cell type-specific genomics of Drosophila neurons. Nucleic Acids Res. 2012; 40: 9691-9704. [CrossRef] [Google scholar] [PubMed]
- Steiner FA, Talbert PB, Kasinathan S, Deal RB, Henikoff S. Cell-type-specific nuclei purification from whole animals for genome-wide expression and chromatin profiling. Genome Res. 2012; 22: 766-777. [CrossRef] [Google scholar] [PubMed]
- Kozlenkov A, Roussos P, Timashpolsky A, Barbu M, Rudchenko S, Bibikova M, et al. Differences in DNA methylation between human neuronal and glial cells are concentrated in enhancers and non-CpG sites. Nucleic Acids Res. 2014; 42: 109-127. [CrossRef] [Google scholar] [PubMed]
- Kozlenkov A, Wang M, Roussos P, Rudchenko S, Barbu M, Bibikova M, et al. Substantial DNA methylation differences between two major neuronal subtypes in human brain. Nucleic Acids Res. 2016; 44: 2593-2612. [CrossRef] [Google scholar] [PubMed]
- Krishnaswami SR, Grindberg RV, Novotny M, Venepally P, Lacar B, Bhutani K, et al. Using single nuclei for RNA-seq to capture the transcriptome of postmortem neurons. Nat Protoc. 2016; 11: 499-524. [CrossRef] [Google scholar] [PubMed]
- Kundakovic M, Jiang Y, Kavanagh DH, Dincer A, Brown L, Pothula V, et al. Practical Guidelines for High-Resolution Epigenomic Profiling of Nucleosomal Histones in Postmortem Human Brain Tissue. Biol Psychiatry. 2017; 81: 162-170. [CrossRef] [Google scholar] [PubMed]
- Sarnat HB, Nochlin D, Born DE. Neuronal nuclear antigen (NeuN): a marker of neuronal maturation in early human fetal nervous system. Brain Dev. 1998; 20: 88-94. [CrossRef] [Google scholar] [PubMed]
- Mullen RJ, Buck CR, Smith AM. NeuN, a neuronal specific nuclear protein in vertebrates. Development. 1992; 116: 201-211. [Google scholar]
- Ransohoff RM, Brown MA. Innate immunity in the central nervous system. J Clin Invest. 2012; 122: 1164-1171. [CrossRef] [Google scholar] [PubMed]
- Gross JA, Pacis A, Chen GG, Barreiro LB, Ernst C, Turecki G. Characterizing 5-hydroxymethylcytosine in human prefrontal cortex at single base resolution. BMC Genomics. 2015; 16: 672. [CrossRef] [Google scholar] [PubMed]
- Shi DQ, Ali I, Tang J, Yang WC. New Insights into 5hmC DNA Modification: Generation, Distribution and Function. Front Genet. 2017; 8: 100. [CrossRef] [Google scholar] [PubMed]
- Huang Y, Pastor WA, Shen Y, Tahiliani M, Liu DR, Rao A. The behaviour of 5-hydroxymethylcytosine in bisulfite sequencing. PLoS One. 2010; 5: e8888. [CrossRef] [Google scholar] [PubMed]
- Booth MJ, Branco MR, Ficz G, Oxley D, Krueger F, Reik W, et al. Quantitative sequencing of 5-methylcytosine and 5-hydroxymethylcytosine at single-base resolution. Science. 2012; 336: 934-937. [CrossRef] [Google scholar] [PubMed]
- Yu M, Hon GC, Szulwach KE, Song CX, Zhang L, Kim A, et al. Base-resolution analysis of 5-hydroxymethylcytosine in the mammalian genome. Cell. 2012; 149: 1368-1380. [CrossRef] [Google scholar] [PubMed]
- Sun Z, Terragni J, Borgaro JG, Liu Y, Yu L, Guan S, et al. High-resolution enzymatic mapping of genomic 5-hydroxymethylcytosine in mouse embryonic stem cells. Cell Rep. 2013; 3: 567-576. [CrossRef] [Google scholar] [PubMed]
- Jones PA. Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat Rev Genet. 2012; 13: 484-492. [CrossRef] [Google scholar] [PubMed]
- Mellen M, Ayata P, Dewell S, Kriaucionis S, Heintz N. MeCP2 binds to 5hmC enriched within active genes and accessible chromatin in the nervous system. Cell. 2012; 151: 1417-1430. [CrossRef] [Google scholar] [PubMed]
- Coppieters N, Dieriks BV, Lill C, Faull RL, Curtis MA, Dragunow M. Global changes in DNA methylation and hydroxymethylation in Alzheimer's disease human brain. Neurobiol Aging. 2014; 35: 1334-1344. [CrossRef] [Google scholar] [PubMed]
- Villar-Menendez I, Blanch M, Tyebji S, Pereira-Veiga T, Albasanz JL, Martin M, et al. Increased 5-methylcytosine and decreased 5-hydroxymethylcytosine levels are associated with reduced striatal A2AR levels in Huntington's disease. Neuromolecular Med. 2013; 15: 295-309. [CrossRef] [Google scholar] [PubMed]
- Dong E, Gavin DP, Chen Y, Davis J. Upregulation of TET1 and downregulation of APOBEC3A and APOBEC3C in the parietal cortex of psychotic patients. Transl Psychiatry. 2012; 2: e159. [CrossRef] [Google scholar] [PubMed]
- Matrisciano F, Tueting P, Dalal I, Kadriu B, Grayson DR, Davis JM, et al. Epigenetic modifications of GABAergic interneurons are associated with the schizophrenia-like phenotype induced by prenatal stress in mice. Neuropharmacology. 2013; 68: 184-194. [CrossRef] [Google scholar] [PubMed]
- Tseng PT, Lin PY, Lee Y, Hung CF, Lung FW, Chen CS, et al. Age-associated decrease in global DNA methylation in patients with major depression. Neuropsychiatr Dis Treat. 2014; 10: 2105-2114. [Google scholar]
- Oved K, Morag A, Pasmanik-Chor M, Oron-Karni V, Shomron N, Rehavi M, et al. Genome-wide miRNA expression profiling of human lymphoblastoid cell lines identifies tentative SSRI antidepressant response biomarkers. Pharmacogenomics. 2012; 13: 1129-1139. [CrossRef] [Google scholar] [PubMed]
- Serafini G, Pompili M, Hansen KF, Obrietan K, Dwivedi Y, Amore M, et al. MicroRNAs: fundamental regulators of gene expression in major affective disorders and suicidal behavior? Front Cell Neurosci. 2013; 7: 208. [CrossRef] [Google scholar] [PubMed]
- Perkins DO, Jeffries CD, Jarskog LF, Thomson JM, Woods K, Newman MA, et al. microRNA expression in the prefrontal cortex of individuals with schizophrenia and schizoaffective disorder. Genome Biol. 2007; 8: R27. [CrossRef] [Google scholar] [PubMed]
- Beveridge NJ, Gardiner E, Carroll AP, Tooney PA, Cairns MJ. Schizophrenia is associated with an increase in cortical microRNA biogenesis. Mol Psychiatry. 2010; 15: 1176-1189. [CrossRef] [Google scholar] [PubMed]
- Belzeaux R, Bergon A, Jeanjean V, Loriod B, Formisano-Treziny C, Verrier L, et al. Responder and nonresponder patients exhibit different peripheral transcriptional signatures during major depressive episode. Transl Psychiatry. 2012; 2: e185. [CrossRef] [Google scholar] [PubMed]
- Bocchio-Chiavetto L, Maffioletti E, Bettinsoli P, Giovannini C, Bignotti S, Tardito D, et al. Blood microRNA changes in depressed patients during antidepressant treatment. Eur Neuropsychopharmacol. 2013; 23: 602-611. [CrossRef] [Google scholar] [PubMed]
- Hannon E, Lunnon K, Schalkwyk L, Mill J. Interindividual methylomic variation across blood, cortex, and cerebellum: implications for epigenetic studies of neurological and neuropsychiatric phenotypes. Epigenetics. 2015; 10: 1024-1032. [CrossRef] [Google scholar] [PubMed]
- Davies MN, Volta M, Pidsley R, Lunnon K, Dixit A, Lovestone S, et al. Functional annotation of the human brain methylome identifies tissue-specific epigenetic variation across brain and blood. Genome Biol. 2012; 13: R43. [CrossRef] [Google scholar] [PubMed]
- Burgess S, Timpson NJ, Ebrahim S, Davey Smith G. Mendelian randomization: where are we now and where are we going? Int J Epidemiol. 2015; 44: 379-388. [CrossRef] [Google scholar] [PubMed]
- Smith GD, Ebrahim S. 'Mendelian randomization': can genetic epidemiology contribute to understanding environmental determinants of disease? Int J Epidemiol. 2003; 32: 1-22. [CrossRef] [Google scholar] [PubMed]
- Relton CL, Davey Smith G. Two-step epigenetic Mendelian randomization: a strategy for establishing the causal role of epigenetic processes in pathways to disease. Int J Epidemiol. 2012; 41: 161-176. [CrossRef] [Google scholar] [PubMed]
- Jhun MA, Smith JA, Ware EB, Kardia SLR, Mosley TH, Jr., Turner ST, et al. Modeling the Causal Role of DNA Methylation in the Association Between Cigarette Smoking and Inflammation in African Americans: A 2-Step Epigenetic Mendelian Randomization Study. Am J Epidemiol. 2017; 186: 1149-1158. [CrossRef] [Google scholar] [PubMed]
- Hynd MR, Lewohl JM, Scott HL, Dodd PR. Biochemical and molecular studies using human autopsy brain tissue. J Neurochem. 2003; 85: 543-562. [CrossRef] [Google scholar] [PubMed]
- Huang HS, Matevossian A, Jiang Y, Akbarian S. Chromatin immunoprecipitation in postmortem brain. J Neurosci Methods. 2006; 156: 284-292. [CrossRef] [Google scholar] [PubMed]
- Stan AD, Ghose S, Gao XM, Roberts RC, Lewis-Amezcua K, Hatanpaa KJ, et al. Human postmortem tissue: what quality markers matter? Brain Res. 2006; 1123: 1-11. [CrossRef] [Google scholar] [PubMed]
- Nagy C, Maheu M, Lopez JP, Vaillancourt K, Cruceanu C, Gross JA, et al. Effects of postmortem interval on biomolecule integrity in the brain. J Neuropathol Exp Neurol. 2015; 74: 459-469. [CrossRef] [Google scholar] [PubMed]
- Stone M, Laughren T, Jones ML, Levenson M, Holland PC, Hughes A, et al. Risk of suicidality in clinical trials of antidepressants in adults: analysis of proprietary data submitted to US Food and Drug Administration. BMJ. 2009; 339: b2880. [CrossRef] [Google scholar] [PubMed]

