

Neurobiology of Sleep and Microbiomics in Aging
Linda J. Larson-Prior 1,2,3,†,*![]() , Gohar Azhar 4
, Gohar Azhar 4![]() , David G. Davila 5
, David G. Davila 5![]() , Se-Ran Jun 6
, Se-Ran Jun 6![]() , Aaron S. Kemp 1
, Aaron S. Kemp 1![]() , Intawat Nookaew 6
, Intawat Nookaew 6![]() , Jeanne Y. Wei 4,7
, Jeanne Y. Wei 4,7![]() , Trudy M. Wassenaar 8,†
, Trudy M. Wassenaar 8,†![]()
-
Department of Psychiatry, University of Arkansas for Medical Sciences, 4801 W. Markham St, Little Rock, AR 72205, USA
-
Department of Neurobiology & Developmental Sciences, University of Arkansas for Medical Sciences, 4801 W. Markham St, Little Rock, AR 72205, USA
-
Department of Neurology, University of Arkansas for Medical Sciences, 4801 W. Markham St, Little Rock, AR 72205, USA
-
Donald W. Reynolds Department of Geriatrics, University of Arkansas for Medical Sciences, 613 Elm St., Little Rock, AR 72205, USA
-
Central Arkansas Veterans Healthcare System Sleep Center, 4300 W. 7th St., 111SL/LR, Little Rock, AR 72205, USA
-
Department of Biomedical Informatics, University of Arkansas for Medical Sciences, 501 Jack Stephens Dr., Little Rock, AR 72205, USA
-
Donald W. Reynolds Institute on Aging, University of Arkansas for Medical Sciences, 613 Elm St., Little Rock, AR 72205, USA
-
Molecular Microbiology and Genomics Consultants, 55576 Zotzenheim, Germany
† These authors contributed equally to this work.
* Correspondence: Linda J. Larson-Prior ![]() .
.
Received: April 5, 2017 | Accepted: April 11, 2017 | Published: April 12, 2017
OBM Neurobiology 2017, Volume 1, Issue 2, doi:10.21926/obm.neurobiol.1702003
Academic Editor: Hermes H. Yeh
Recommended citation: Larson-Prior LJ, Azhar G, Davila DG, Jun SR, Kemp AS, Nookaew I, Wei JY, Wassenaar TM. Neurobiology of Sleep and Microbiomics in Aging. OBM Neurobiology 2017; 1(2): 003; doi:10.21926/obm.neurobiol.1702003.
© 2017 by the authors. This is an open access article distributed under the conditions of the Creative Commons by Attribution License, which permits unrestricted use, distribution, and reproduction in any medium or format, provided the original work is correctly cited.
Abstract
(1) Background: The daily rhythms of life on earth are governed by a central clock located in the hypothalamus and entrained primarily by light. By virtue of its connections to a myriad of other clocks located both in the brain and in peripheral organs and cells, this clock provides a timing signal based on the light/dark cycle by which organismal physiology and behaviour are governed. Disturbances of these natural rhythms have multiple downstream effects, frequently resulting in dysregulations of sleep and wake cycles, energy homeostatic signalling and decreased stability of the gut microbiome. With increasing age, changes in these natural rhythms are common, and distinguishing changes associated with healthy aging from those associated with pathological aging is an important and growing avenue of investigation in which the recognition of the interdependence of these systems is critical. (2) Aims: In this review, we focus on the importance of better understanding the integration of these signals and their physiological and behavioural outputs in an aging population in which sleep durations are frequently shortened and dietary choices highly varied and frequently nutritionally poor. (3) Conclusions: The gut-brain-microbiome axis is a critical regulator of health across the lifespan such that dysregulation at any point along this axis impacts organismal function. As yet, the impact of age alone on this system is not fully understood. However, it is clear that age-related dysfunctions including the development of frailty and the increased risk for neurological disease are intertwined with the health of the gut-brain-microbiome axis.
Keywords
Sleep; Energy homeostasis; Gut microbiome; Aging; Gut brain axis; circadian rhythm
Introduction
Life on earth is organized rhythmically; from the rhythmic alteration of the seasons to the cycling of day and night. From the organismal level down to the clock-like rhythms generated by single cells and cellular networks, timing is important. These rhythms pervade the lives of all organisms and, when disrupted, can lead to adverse consequences ranging from increased desire for sleep to significantly reduced physical and mental health, or even, in the extreme case, to premature death.
In mammals, daily rhythms are tied to exposure to light and dark. These circadian (from ‘circa’ approximately, and ‘dies’ day) cycles follow the intrinsically generated rhythms of a central clock located in the suprachiasmatic nucleus (SCN) of the hypothalamus (Figure 1). The timing of this central clock, known as the circadian oscillator (CO), is synchronised by light (intensity, wavelength, timing and duration), sensed by the retina and signalled to the brain via the retino-hypothalamic tract [1]. The ‘default setting’ of the SCN clock is slightly more than 24 hours, which is reset to match the Earth’s day and night by exposure to light, in particular to morning light [2]. Apart from this input, the SCN clock is fine-tuned (or disturbed) by regular feeding/fasting patterns, and to a lesser extent by temperature fluctuations and other external factors.
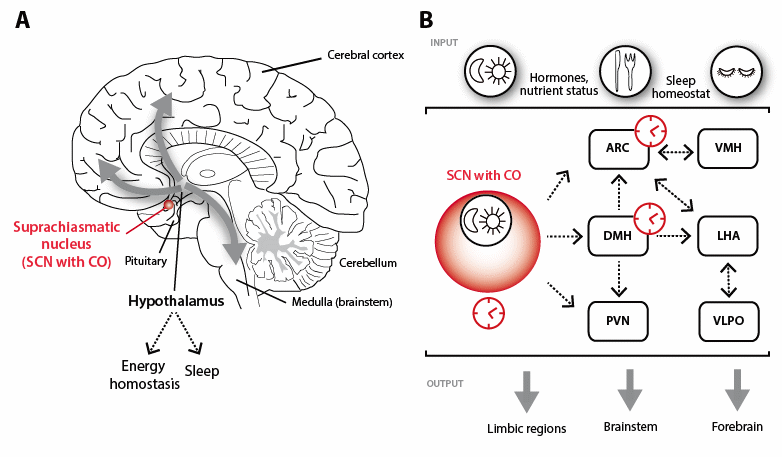
Figure 1 Hypothalamic control of sleep and feeding. (A) The suprachiasmatic nucleus (SCN, indicated in red) contains the circadian oscillator (CO) and provides signals to hypothalamic nuclei involved in the regulation of sleep/wake rhythms and energy homeostasis. (B) Hypothalamic nuclei under control of the SCN include the arcuate nucleus (ARC) and ventromedial hypothalamus (VMH) as primary feeding centers. The lateral hypothalamic area (LHA) and the ventrolateral preoptic nucleus (VLPO) comprise regulatory centers for wake and sleep respectively and receive indirect SCN input via the dorsomedial hypothalamic nucleus (DMH). The DMH provides a central integrating center for many biological rhythms, including those of sleep/wake and energy homeostasis, as does the LHA. The paraventricular nucleus (PVN) also receives direct SCN input and provides signalling important in sleep and feeding behaviours via both sympathetic and parasympathetic afferent limbs. Inputs to the system include hormonal, peptidergic and neural information on nutrient status, energy utilization and sleep need while system outputs link to higher order cognitive centers, limbic centers and brainstem as well as to peripheral organ systems.
This central clock provides timing information to a large network of clocks that are found in peripheral organs, tissues and cells that are themselves able to generate autonomous rhythms [3,4]. While many models of this system posit a hierarchical system in which the SCN provides the primary clock signal to which peripheral clocks are tightly entrained [5], recent evidence suggests a far more complicated network in which peripheral organs may independently entrain to external signals [4] and provide feedback information to both SCN and other peripheral clocks [3]. An overview of this circuit, which links sleep/wake and fasting/feeding rhythms in the hypothalamic circuitry, is illustrated in Figure 1B. Briefly, the SCN provides direct timing signals to the arcuate nucleus (ARC), a central regulator of energy homeostasis, the dorsomedial hypothalamic nucleus (DMH) and the paraventricular nucleus (PVN). The SCN further provides indirect timing via the DMH to central regulating centers for wake and feeding (lateral hypothalamic area, LHA) and sleep (ventrolateral preoptic area, VLPO). Both ARC and DMH express their own molecular clocks, and the DMH acts to integrate circadian clock timing with neuroendocrine (via PVN), sleep (via LHA) and feeding (ARC and LHA) behaviours among others [6]. The role of ARC in energy regulation depends in large part on the balance between its two primary neuronal populations; those cells co-expressing neuropeptide-Y (NPY), agouti-related protein (AgRP) and GABA and those co-expressing pro-opiomelanocortin (POMC) and cocaine-and-amphetamine-regulated transcript (CART). Together with the melanocortin receptors and their target systems, these comprise the melanocortin system, which is critical in energy homeostasis [7,8]. The role of the LHA in wake and feeding is largely through neuronal systems expressing orexin/hypocretin (ORX) and melanin-concentrating hormone (MCH), with the ORX system playing a major role in both wake and feeding, and MCH-containing neurons acting to facilitate sleep [9,10]. Thus, the LHA and DMH are important integrators of the complex interplay of sleep, feeding and energy homeostasis in organismal function [8,10,11].
The daily routines of waking and sleeping, feeding and fasting regulate basic physiological processes of the body, including the digestive system and metabolism, the activity of the immune system and the reproductive system, the cardiovascular and endocrine systems, and various brain regions. By virtue of this complex network, the biological machinery of life resonates to the rhythms of the external world. This review is focused on the interdependent rhythms that tie the gut-brain axis and sleep-wake cycles and how they are altered during the aging process under the premise that, to promote the healthspan, biomedicine must look beyond single physiological systems to better understand the interactions and dependencies among them.
General Patterns of Sleep, Wake and Feeding
Embedded in the circadian rhythm are the shorter (ultradian) rhythms of sleep. In adult humans, this sleep rhythm is consolidated into the dark period and lasts between four and 10 hours, with an average of 7–8 hours. The sleep period alternates rhythmically with waking, which is consolidated to the light period in normal circumstances (Figure 2).
The sleep period consists of three defined periods that rhythmically alternate in 2–7 cycles over a single sleep bout in normal sleepers [12]. Those periods are defined based upon characteristic waveforms in the scalp-recorded electrical activity of the brain and consist of (1) N1 or sleep onset, (2) Non-rapid eye movement (NREM) sleep which consists of two electrographically distinct periods—light sleep (N2) and deep sleep (N3), from which sleepers are difficult to arouse, and (3) rapid eye movement (REM) sleep, named for the pattern of eye movements characteristic of the period. In the first sleep cycle of the night, there is a general progression from wake to N1, on to N2, followed by N3. There is frequently a brief arousal or movement as N3 transitions to REM sleep in this first cycle. In subsequent cycles, this pattern persists although sleepers no longer transition through the N1 stage, instead moving from REM directly into N2 to repeat the pattern. Over the course of the normal sleep period, the amount of time spent in N3 is greatest early in the cycle and decreases over the night, while REM sleep exhibits the opposite pattern and is of longest duration in the later cycles.
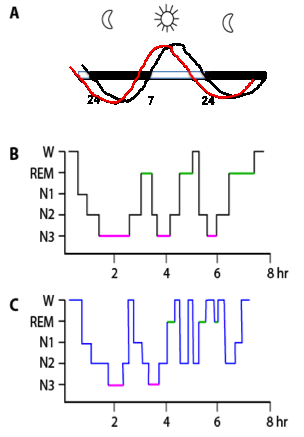
Figure 2 Sleep patterns in adults and elderly. (A) Diagram of the circadian timing of the drive for arousal in adults (black curve) and aging individuals (red curve). (B) Sleep histogram over a night of sleep in normal young adulthood. Note the prominence of N3 (also known as slow wave sleep, SWS, pink line) early in the night and the progressive increase in REM (green line) late in the sleep period. (C) Sleep histogram over a night’s sleep in aging, with shorter overall sleep duration due to frequent arousals that result in the fragmentation of sleep and reductions in both N3 and REM durations.
The most widely accepted model of the relationship between circadian and sleep rhythms is known as the two-process model, and was first proposed by Borbely [13]. In this model, the circadian system comprises one arm (Process C) while the sleep homeostat comprises the second (Process S). The circadian arm of this model promotes preferred periods of wake and sleep, while the sleep homeostat tracks prior sleep history to control the need for sleep. A mismatch in timing between these two processes, such as occurs with jet lag, results in poor daytime function and disturbed sleep until the two rhythms become realigned. While originally posited to be independent processes, more recent work indicates that these two processes are integrated at the molecular level [14,15,16].
In concert with the daily rhythms of waking and sleeping are those of feeding and fasting, essential components of the energy regulation system of the organism. Feeding behaviour relies on the interactions of metabolic, endocrine, autonomic and cognitive factors and is generally timed for the active period [8,17]. At its core, energy homeostasis depends on the regulation of both energy intake via feeding and energy expenditure, and many of the organ systems associated with these processes exhibit their own clock gene rhythms including the liver, pancreas, gastrointestinal tract, adipose tissue and skeletal muscle [8,18,19]. Each of these peripheral organs produces metabolic signals that display circadian rhythmicity, including glucose (liver), insulin (pancreas) and leptin (adipose tissue), and disruptions of these rhythms have been shown to result in impaired energy regulation [18,19]. To maintain energy balance, the intake of nutrients must be balanced by energy expenditures that include basal metabolic rates, thermogenesis and physical activity [20]. Thus, areas involved in energy homeostasis must receive information on the status of energy intake and storage as well as on energy usage and expenditure. To accomplish this, peripheral and central oscillators are regulated not only by the SCN, but also by nutrients that become available after a meal, and hormonal signals from both peripheral and central energy sensors. The effect of nutrients on these oscillators may be even stronger than that of the SCN, enabling shifts in cycle times that no longer match the circadian clock output timing. For example, whereas blood glucose levels are hormonally regulated by insulin and glucagon activity, whose production is again under regulation of the circadian clock (insulin blood levels typically peak around 1700 hr and are lowest around 0400 hr), these levels also respond to food intake, overriding the general background regulation. As a result, feeding at times not in phase with the circadian clock can upset synchronization [21].
Recent work has revealed the presence of an apparent clock for feeding that is termed the food entrainable oscillator (FEO) [6,22]. The existence of this oscillator was signalled by studies in which animals were observed to increase locomotor activity 2–3 hours prior to an established mealtime, a phenomenon that is known as food-anticipatory activity (FAA) [6,22]. Later studies, in which a restricted feeding schedule was introduced with feeding times shifted relative to the circadian clock, showed that FAA entrained to this shifted time and the timing of the sleep-wake cycle shifted with it, so that animals awoke a few hours before the re-scheduled feeding time [6,19,22,23]. Interestingly, a recent study in rodents reported that clear shifts in sleep architecture entrained to the FEO under a food entrainment protocol, indicating a close coupling between feeding and sleep rhythms [23]. Despite studies designed to locate the FEO, no clear region appears to generate this rhythm, which is currently speculated to be generated by a network of oscillators outside the light-entrained hypothalamic clock [6,23].
Although the body is well equipped to adjust sleep and feeding patterns to the phases of day and night, modern lifestyles and exposure to artificial light has led to a continuous override of this rhythm, resulting on average in less sleep than was common in the pre-electricity era [24]. Chronic sleep reduction results in poorer health outcomes, including an increased risk of obesity and metabolic syndrome that appear largely due to increases in food intake [25,26,27,28]. Shift work, which represents a misalignment of the circadian clock and the day/night cycle, is also associated with significant health risks, including obesity [29,30]. These data point to the importance of considering the interactions between sleep, energy homeostasis and the circadian timing cycle in health and disease.
Sleep, Wake and Feeding in Aging
The patterns of both sleeping and feeding change with increasing age, in part due to shifts in the output of the circadian oscillator of the SCN. Although aging is accompanied by changes in most rest-activity oscillations, the neurons of the SCN maintain their clock function with aging [31,32]. Thus, the age-related changes in clock function are due to deterioration of SCN network function that dampens the clock signal passed to downstream oscillators [31,32,33,34]. In addition, there is a shift in circadian timing, expressed largely in preferred sleep and wake timing, with a phase advance leading to early preferred sleep times and wake times (Figure 2A) [35,36]. Significant changes in the patterning and duration of nocturnal sleep are also seen with increasing age [32,36,37]; including reductions in total sleep duration, increased sleep latency, increased waking during the night leading to fragmented sleep and reduced sleep efficiency (Figure 2C). These changes in sleep duration and patterning are likely to underlie changes in cognitive function that can accompany both normal and pathological aging [38,39]. In addition, the ability to accommodate sudden shifts in the circadian phase, such as those introduced by jet lag or shift work, decreases with increased age and may be related to a blunted response of the SCN to light [34,35]. Although sleep disturbances are common, they are not ubiquitous. For many older adults medical, psychiatric or psychosocial factors also contribute to age-related changes in sleep patterns that together can result in increased morbidity and mortality [36].
In general, aging is associated with a reduction in physical activity and skeletal muscle mass, resulting in a decline in energy requirements [19] that often corresponds to a decrease in caloric intake [40,41]. While there is a recent trend among older adults toward increasing levels of obesity due to increased ingestion of high caloric value foods [19,42], the more common pattern has been a reduction in caloric intake with aging. A frequent concomitant to this pattern is an extreme reduction in food intake that leads to caloric intake insufficient to meet energy needs [40,41,43] termed anorexia of aging. Physiological changes such as reductions in response to orexigenic hormones like ghrelin and cholecystokinin have been suggested to play a role, as have reductions in hedonic aspects of feeding, but it is clear that social and environmental factors also play a major role in the loss of appetite that can be exacerbated by disease, economic status and mental health [40,42,43]. Among these, heart failure affects over 10% of the older population and is the leading cause of morbidity and mortality in this demographic. Advanced heart failure results in unintentional weight loss, muscle weakness and a syndrome of cardiac cachexia. Right heart failure in particular directly affects the gut by causing mucosal edema leading to gut ischemia (leaky gut syndrome) with disturbances of nutrient absorption and abnormal bacterial translocation [44,45]. The outcome of this feeding pattern is reduced mobility and physical activity levels, either sarcopenic obesity or frailty, a worsening nutritional status and an increased risk of all-cause mortality [41,42,43,46,47].
With aging, the biological processes controlling energy consumption and utilization become less efficient, leading to increased damage to the cells and tissues engaged in these critical functions [48,49]. The aging process extends to core metabolic functions as well, including glycolysis, fatty acid and amino acid oxidation, and lipogenesis [48,49,50]. Peripheral clocks located in the liver and pancreas that regulate metabolic rhythms such as glucose homeostasis and lipid metabolism also exhibit reduced amplitude with advancing age [35]. Overall, metabolic flexibility—the ability of cells and tissues to switch between metabolic functions of the fasting state to those involved in energy storage following feeding—is believed to be significantly reduced in older adults [50,51].
In addition, nutritional state and sleep status affect metabolic and immune system functions such that aging is often accompanied by a gradual development of low-grade chronic systemic inflammation that increases the risk of age-associated disease [10,51,52,53,54,55,56]. Diet and feeding modulate immune function both directly and indirectly via actions on the gut microbiome, which is a key regulator of both immune and metabolic function [55,56,57]. Sleep restriction, whether alone or in combination with misalignment of circadian rhythms (e.g., shift work) often results in reduced sleep quality and dysregulated glucose control [54,55]. Chronic reductions in sleep, whether from volitional changes in sleep patterning or age-related disruptions, lead to alterations in energy intake and expenditure, unstable weight regulation and an increase in pro-inflammatory cytokine levels [26,54,55,58]. Together, these data indicate the closeness of interactions between sleep, energy homeostasis, and the gut microbiome that can become dysregulated with advancing age and lead to increased risk for multi-system failure.
The Gut Microbiome in Aging
Bacterial communities are found on all surfaces in contact with the outside world. Not only skin and hair, but also the mucosal surface of oral and nasal cavities, airways, gastro-intestinal tract, and uro-genital tract are covered by bacteria, and to a lesser extent by archaea, viruses, fungi and other monocellular organisms. All these different body surfaces support the growth of microbial communities (microbiomes) of different, site-specific compositions [59]. Due to the next generation sequencing revolution, the gut microbiome is the best described, exhibiting a clear individual signature, though there is known variation over time and at different locations along the gut [60,61]. Despite some day-to-day variation in the presence or absence of individual bacterial species, the gut microbiome of an individual is roughly constant, as long as its composition is not disturbed by internal or external factors such as disease, changes in diet, or intake of antibiotics [59,60,61]. When comparing human populations, cultural differences can be reflected by differences in the gut microbiome, as when populations from the US or Europe are compared to rural Africa or South-America [62,63]. On an even larger scale, the human microbiome can be recognized and differentiated from that of other mammals, particularly those with different feeding patterns (ruminants, carnivores). At the individual level, the microbiome of siblings is more similar than that of non-related individuals [63,64]. A healthy gut microbiome (in which the majority of bacteria will be located in the colon) contains three prominent bacterial phyla: the Bacteroides, Firmicultes and Proteobacteria [65,66]. The composition of the gut microbiota changes with feeding as a function of the species-specific circadian cycle such that it functions in energy metabolism during the active phase and detoxification processes in rest phases [55].
The human gut microbiome functions to extend the human genome by adding a variety of bioactive substances that are not coded for in the human genome. These function in the extraction of energy from ingested meals, fermentation of fiber, production of short-chain fatty acids, modulation of the immune system and vitamin synthesis [66,67]. The numerous variations in physical activity, dietary habits and cultural norms concerning feeding times and dietary composition have made it difficult to definitively identify the composition of a healthy microbiome, despite an increasing number of studies pointing to changes in the gut microbiome in disease [65,68,69], sleep restriction [28,70], inflammation [68,71], sedentary behaviours [72], and dietary composition [61,69].
While a clear definition of the ‘healthy aged gut’ remains elusive, age-related spatio-temporal variations in the gut microbiota such as a decrease in Bifidobacteria and Firmicutes together with a decrease in overall microbiota diversity are commonly reported [61,73,74]. Age-related disruption of the gut microbiome plays an important role in pathological states associated with aging such as frailty [74]. As noted in the previous discussion of anorexia of aging, development of frailty in older adults is associated with impaired physiological functioning, reduced resistance to stressors and adverse health outcomes. A recent study examining changes in the gut microbiome associated with frailty in community dwelling female twins reports a significant reduction in diversity together with changes in the abundance of specific taxa [75] although the directionality of these changes could not be assessed in this study. It is also the case that certain variants of the gut microbiome are associated with increased longevity, where the microbial composition is characterised by an increase in anti-inflammatory species and a loss of genes involved in short-chain fatty acid synthesis [74]. Thus, while the microbiome in healthy aging as defined by an extended lifespan shows changes consistent with that of shorter lived elders, there are compensatory increases in taxa that were subdominant in adulthood that appear to provide health benefits, allowing us to envision microbiota-based health surveillance for older adults as an innovative strategy.
In the developing organism, gut bacteria play an important role in shaping the gut-associated lymphatic tissue (GALT) and training the immune system in a critical bilateral communication system between the host immune system and its colonizing microorganisms [76,77]. With advancing age, the functionality of the immune system often declines, resulting in a chronic low-grade inflammatory status in the gut that coincides with changes in gut microbiome composition and stability [57,78]. As noted previously, reductions in sleep duration and quality associated with older age can act to further exacerbate reductions in immune system function [26,55]. An increase in the abundance of gram-negative bacteria in the aging gut has been suggested to lead to secretion of lipopolysaccharides that can result in inflammation and leakiness of the gut, especially when coupled with a reduction in the level of short chain fatty acids such as butyrate in the intestine that may increase intestinal porosity [61].
The Gut-Brain-Microbiome Axis
The relationship between human beings and their microbiota is symbiotic, with those microorganisms that act to promote overall health considered symbionts and those that induce pathological responses termed pathobionts [68]. The concept of the gut-brain axis recognizes the integrative functioning of the nervous and gastrointestinal systems, incorporating the neural afferent and efferent signals together with endocrine, nutrient and immunological signals that form the bidirectional communication portals between these organ systems [79]. In addition to its role in gut function, immune activation and energy homeostasis, the gut-brain axis links gastrointestinal function to emotional and cognitive centers of the brain [80], with an increasing number of studies indicating a significant role of the gut-brain axis in human pathologies, ranging from Irritable Bowel Disease to anxiety disorders [69,76,81,82].
The bidirectional communication network of the gut-brain axis includes components of the CNS (brain and spinal cord), autonomic nervous system (ANS), enteric nervous system (ENS), the intestinal microbiota and the hypothalamic pituitary adrenal (HPA) axis as well as hormonal signalling elements from the gut lumen [80,83,84]. The vagus nerve provides innervation to the muscular and mucosal layers of the gut, providing information on immune, microbial and nutrient information to the CNS, while stimulation of the efferent limb of the vagus nerve results in the release of pro-inflammatory cytokines [85]. While the vagus nerve provides an important conduit in the bidirectional information flow in the gut-brain axis, extra-vagal systems also play major roles.
The enteric nervous system, though in communication with the CNS via sympathetic and parasympathetic arms of the ANS, is an autonomous nervous system that controls the function of gastrointestinal tract (GIT). The ENS consists of a mesh-like system of neurons and fibers that encase the GIT from the level of the upper esophagus to the internal anal sphincter [86,87]. The ENS network is made up of primary sensory neurons, interneurons and motor neurons that control GIT motility, digestive enzyme secretions, fluid exchange, barrier function, immune responses and blood flow [88]. The ENS develops in concert with the enteric microenvironment, which thus exerts some control over the neuronal composition and function of this system based on variations in diet, microbial composition, disease, inflammation and medication [78,86]. The ENS neurons also utilize over 30 neuroactive substances that are similar to those of the CNS, including acetylcholine, serotonin, catecholamines (epinephrine, norepinephrine and dopamine) and gamma-butyric acid (GABA) [76,78,85,87,88,89]. There is direct hormonal communication between the gut-brain axis and the HPA axis, the primary stress response system, a relationship critical to normal functioning of both systems [68,78,90]. Stress exposure affects the gut microbiota and activates the HPA axis, which together can increase the risk of inflammation-related diseases, and decrease the threshold for anxiety [69,86,90].
A number of studies have examined the role of the microbiota in behaviour and disease, finding intriguing connections that link the microbiome-gut-brain axis to both immune-related and non-immune related diseases including multiple sclerosis, anxiety and depression, autism spectrum disorders and Parkinson’s Disease (PD) [69,82]. For the purposes of this review, PD represents a particularly intriguing linkage between sleep, energy homeostasis and the microbiome as prodromal symptoms now recognized to occur decades or years before the onset of clinical symptoms include autonomic dysfunctions, constipation, and sleep disorders. Further, according to the widely accepted Braak hypothesis of disease progression, early markers of disease onset occur in the olfactory bulbs and ENS, where the vagal nerve may act to facilitate the spread of alpha synuclein to medullary centers that are known to be affected early in the disease progress [82,91]. A role for the gut-brain axis in PD is suggested by studies showing a reduced risk of PD following truncal vagotomy and a reduced risk of PD-related pathophysiology in germ-free mice overexpressing α-synuclein [82].
Dysbiosis, Aging and Desynchronized Cycles
The term dysbiosis (also spelled disbiosis) was coined to describe a situation in which the gut microbiome is unbalanced in composition, thus deviating from what can be considered normal. The situation is common in the elderly, for a number of reasons, not in the least due to frequent antibiotic use. The longer individuals live in long-stay care facilities, the more gut bacterial diversity reduces, which results in a less resilient microbiome [92]. Enteric infections are more common in the elderly, in whom higher levels of Clostridium difficile may be present [93]. An early, polyphasic (though culture-dependent) study compared the faecal microbiota of elders to that of younger adults and children, identifying a decrease in Bifidobacteria species in the elderly [94]. These authors not only mention antibiotic intake and altered immune responses, but also decreased gastric acid secretion and increased gut mucosal permeability as underlying causes for dysbiosis. Antibiotic intake may be required for gastro-enteric and other infections (airways, urinary tract infections, etc.), resulting in reduced microbiome diversity [95]. Age-associated alterations in feeding patterns also occur in common co-morbid conditions of the elderly such as chronic heart failure, renal failure, chronic obstructive lung disease and cancer, all conditions that lead to weight loss, changes in immunity and an abnormal gut-microbiota muscle axis, with sarcopenia and progressive weakness [96]. This can create a vicious circle, as a less diverse microbiome is more susceptible to enteric pathogens. These and other factors affecting the gut microbiome are schematically shown in Figure 3A. For the elder population, the most frequent factors that influence the gut microbiome towards dysbiosis and an unhealthy status are antimicrobial intake, changes in diet and food habits, changes in immune function, and changes in sleeping patterns.
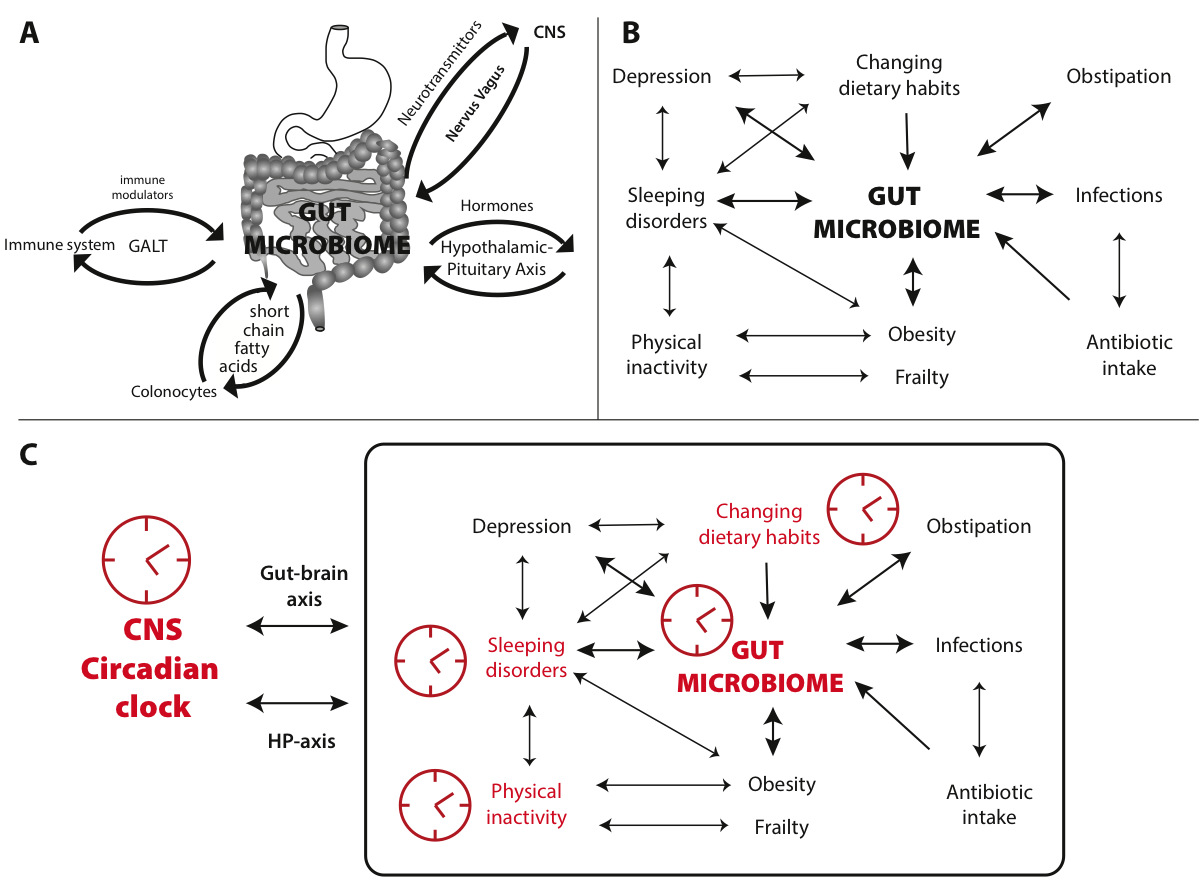
Figure 3 Interplay of a dysbiose, aging and circadian cycles. (A) Schematic of the bidirectional interactions between the gut microbiome and the immune system (via immune modulators communicating with GALT), the endocrine system (via hormones acting through the hypothalamic-pituitary axis), the central nervous system (via neurotransmittors traveling through the vagal nerve) and colonocytes (via short-chain fatty acids). (B) Illustrates factors frequently affecting the gut microbiome of the elderly, indicating their mutual enforcement. (C) Combining the effect of these interactions with circadian cycles (symbolized by the clocks), with major effects shown in red, shows how desynchronization can disturb homeostasis and adversely impact physical and mental health.
Several of these factors enforce each other in feedback loops, schematically shown in Figure 3B. It is therefore no surprise that specific interventions affecting the gut microbiome are now considered for treatment of dysbiosis and its related diseases, in particular to improve health in the elderly [74]. After all, many of the chronic diseases associated with shifts in the gut microbiome, and by some considered to be the result of these shifts, are typically observed with increasing age, such as obesity, insulin resistance, type 2 diabetes, metabolic syndrome, Inflammatory Bowel Disease and non-alcoholic fatty liver diseases [97]. Microbiota-targeted interventions, such as intake of probiotics (live bacteria which confer a health benefit on the host) or prebiotics (nonviable food components that confer a health benefit on the host associated with modulation of the microbiota) are now becoming mainstream [98]. Although such interventions are considered by some as promising anti-aging options, cause and effect in the gut microbiome and associated pathologies is often not clear. In view of the intricate communication pathways between the gut microbiome and the host, this is not surprising. Understanding these complex systems requires a holistic [99] rather than a reductionist approach; it does not suffice to study simplified models in which the effects of one bacterial species only are examined. In view of the wide variety of microbiome composition between individuals (only one third of the gut bacteria is considered to be shared by most individuals sharing a geographic and cultural background, with two-third being variable), it is probably simplistic to assume significant effects as a result of deliberate intake of a single probiotic bacterial strain; to expect the same beneficial effect in all individuals, or for it to treat a variation of health problems, is even less realistic.
To the complexity of interactions between gut microbiome and host, we can now add the effect of circadian rhythmicity of the gut bacteria [77]. An intimate relationship exists between the gut microbiome and the circadian regulators produced by the host’s intestinal epithelial cells, as was demonstrated in rodents [100]. The result is a circadian rhythm both in microbial activity and in nutrient uptake by the gut epithelial cells, in which Toll-like receptors (TLR) are involved [101]. The resulting diurnal microbial behaviour drives, in turn, the global programming of the host circadian oscillations [77]. Disruption of this microbiome rhythmicity has impact on the host physiology. Just as desynchronization of neural clocks can result in negative health effects, desynchronized circadian cycles of the gut can worsen health status, through the interactions shown in Figure 3C.
Although the literature on this subject is still limited, one can envision that a desynchronization of one or more circadian cycles may affect gut activity, and vice versa. In this respect it may be relevant to note that constipation [102] and depression are relatively common in the elderly, both of which have been linked to shifts in the microbiome. However, the exact role of the microbiome in age-related health issues is still insufficiently explored [79].
Conclusion
The processes in place to maintain system homeostasis and overall health are multiple, complex and interconnected. The neurobiological rhythms that collectively govern a host of cyclical behaviours in humans may have pervasive bidirectional relations with the bacterial communities that human’s host. Understanding how these complex bidirectional interdependencies are altered by daily sleep/wake and feeding cycles and in turn across an individual’s lifespan may prove critical for discovering how these factors influence the balancing of fragility and resilience when confronted with pathological perturbations. Accordingly, research into the neurobiology of sleep, microbiomics, and aging is very likely to be instrumental in developing effective interventions for a wide range of human diseases and disorders.
Author Contributions
Dr. Larson-Prior and Dr. Wassenaar contributed equally to this review. Dr. Azhar, Dr. Davila, Dr. Jun, Mr. Kemp, Dr. Nookaew and Dr. Wei provided valuable input in discussions of the material presented and helped to edit the manuscript.
Competing Interests
The authors declare that no competing interests exist.
References
- Hannibal J. Neurotransmitters of the retino-hypothalamic tract. Cell Tissue Res. 2002;309(1):73-88. PMID: 12111538 [CrossRef] [Google scholar] [PubMed]
- Richardson GS. The human circadian system in normal and disordered sleep. J Clin Psychiatry. 2005;66 Suppl 9:3-9. PMID: 16336035 [Google scholar]
- Menaker M, Murphy ZC, Sellix MT. Central control of peripheral circadian oscillators. Curr Opin Neurobiol. 2013;23:741-6. [CrossRef] [Google scholar] [PubMed]
- Husse J, Eichele G, Oster H. Synchronization of the mammalian circadian timing system: light can control peripheral clocks independently of the SCN clock. Bioessays. 2015;37:1119-28. [CrossRef] [Google scholar] [PubMed]
- Partch CL, Green CB, Takahashi JS. Molecular architecture of the mammalian circadian clock. Trends in Cell Biol. 2014;24(2):90-9. [CrossRef] [Google scholar] [PubMed]
- Bechtold DA, Loudon ASI. Hypothalamic clocks and rhythms in feeding behaviour. TINS. 2013;36:74-82Crone. [Google scholar]
- Crone RD. Anatomy and regulation of the central melanocortin system. Nat Neurosci. 2005;8:571-8. [CrossRef] [Google scholar] [PubMed]
- Caron A, Richard D. Neuronal systems and circuits involved in the control of food intake and adaptive thermogenesis. Ann NY Acad Sci. 2017;13263:35-53. [CrossRef] [Google scholar] [PubMed]
- Monti JM, Torterolo P, Lagos P. Melanin-concentrating hormone control of sleep. Sleep Med Rev. 2013;17:293-8. [CrossRef] [Google scholar] [PubMed]
- Nixon JP, Mavanji V, Butterick TA, Billington CJ, Kotz CM, Teske JA. Sleep disorders, obesity and aging: the role of orexin. Ageing Res Rev. 2015;20:63-73. [CrossRef] [Google scholar] [PubMed]
- Herrera CG, Ponomarenko A, Korotkova T, Burdakov D, Adamantidis A. Sleep & metabolism: The multitasking ability of the lateral hypothalamic inhibitory circuits. Front Neuroendocrin. 2017;44:27-34. [CrossRef] [Google scholar] [PubMed]
- Carskadon MA, Dement WC. Principles and practice of sleep medicine. 5th ed. St. Louis: Elsevier Saunders; 2011. p. 16-26. [CrossRef] [Google scholar]
- Borbely AA. A two process model of sleep regulation. Human Neurobiol. 1982;1(3):195-204. [Google scholar]
- Franken P. A role for clock genes in sleep homeostatsis. Curr Opin Neurobiol. 2013;23:864-72. [CrossRef] [Google scholar] [PubMed]
- Curie T, Mongrain V, Dorsaz S, Mang GM, Emmenegger Y, Franken P. Homeostatic and circadian contribution to EEG and state variables of sleep regulation. Sleep. 2013;36(3):311-23. [CrossRef] [Google scholar] [PubMed]
- Borbely AA, Daan S, Wirz-Justice A, Deboer T. The two-process model of sleep regulation: a reappraisal. J. Sleep Res. 2016;25:131-43. [CrossRef] [Google scholar] [PubMed]
- Willie JT, Chemelli RM, Sinton CM, Yanagisawa M. To eat or sleep? Orexin in the regulation of feeding and wakefulness. Ann Rev Neurosci. 2001;24:429-58. [CrossRef] [Google scholar] [PubMed]
- Huang W, Ramsey KM, Marchva B, Bass J. Circadian rhythms, sleep, and metabolism. J Clin Invest. 2011;121:2133-41. [CrossRef] [Google scholar] [PubMed]
- Johnston JD. Physiological responses to food intake throughout the day. Nutrition Res Rev. 2014;27:107-18. [CrossRef] [Google scholar] [PubMed]
- Waterson MJ, Horvath TL. Neuronal regulation of energy homeostasis: beyond the hypothalamus and feeding. Cell Metab. 2015;22(6):962-70. PMID: 26603190 [CrossRef] [Google scholar] [PubMed]
- Yasumoto Y, Hashimoto C, Nakao R, Yamazaki H, Hiroyama H, Nemoto T, et al. Short-term feeding at the wrong time is sufficient to desynchronize peripheral clocks and induce obesity with hyperphagia, physical inactivity and metabolic disorders in mice. Metabolism. 2016;65(5):714-27. PMID: 27085778 [CrossRef] [Google scholar] [PubMed]
- Carneiro BTS, Araujo JF. Food entrainment: major and recent findings. Chronobiol Int. 2012;26:1273-1289. [CrossRef] [Google scholar] [PubMed]
- Castro-Faundez J, Diaz J, Ocampo-Barces A. Temporal organization of the sleep-wake cycle under food entrainment in the rat. Sleep. 2016;39:1451-1465. [CrossRef] [Google scholar] [PubMed]
- Foster RG, Wulff K. The rhythm of rest and excess. Nat Rev Neurosci. 2005;6(5):407-14. Review. PMID: 15861183 [Google scholar]
- Markwalk RR, Melanson EL, Smith MR, Higgins J, Perreault L, Eckel RH et al. Impact of insufficient sleep on total daily energy expenditure, food intake and weight gain. PNAS. 2013;10:5695-700. [CrossRef] [Google scholar] [PubMed]
- Chaput, J-P. Sleep patterns, diet quality and energy balance. Physiol Behav. 2014;134:86-9. [CrossRef] [Google scholar] [PubMed]
- Grandner MA, Jackson N, Gerstner JR, Knutson KL. Sleep symptoms associated with intake of specific dietary nutrients. J Sleep Res. 2014;23:22-34. [CrossRef] [Google scholar] [PubMed]
- Poroyko, VA, Carreras A, Khalyfa A, Khalyfa AA, Leone V, Peris E et al. Chronic sleep disruption alters gut microbiota, induces systemic and adipose tissue inflammation and insulin resistance in mice. Scientific Reports. 2016;6:35405. [CrossRef] [Google scholar] [PubMed]
- Maywood ES, O'Neill J, Wong GK, Reddy AB, Hastings MH. Circadian timing in health and disease. Prog Brain Res. 2006;153:253-69. PMID: 16876580. [CrossRef] [Google scholar] [PubMed]
- Antunes LC, Levandovski R, Dantas G, Caumo W, Hidalgo MP. Obesity and shift work: chronobiological aspects. Nutrition Res Rev. 2010;23:155-68. [CrossRef] [Google scholar] [PubMed]
- Manoogian EN, Panda S. Circadian rhythms, time-restricted feeding, and healthy aging. Ageing Res Rev. 2016; doi:10.1016/j.arr.2016.12.006. PMID: 28017879 [CrossRef] [Google scholar] [PubMed]
- Mattis J, Sehgal A. Circadian rhythms, sleep, and disorders of aging. Trends in Endorcrinology & Metabolism. 2017;27:192-203. [CrossRef] [Google scholar] [PubMed]
- Monk TH. Aging human circadian rhythms: conventional wisdom may not always be right. J Biol Rhythms. 2005;20(4):366-74. PMID: 16077155 [CrossRef] [Google scholar] [PubMed]
- Liu F, Chang H-C. Physiological links of circadian clock and biological clock of aging. Protein Cell. 2017; doi:10.1007/s13238-016-0366-2. [CrossRef] [Google scholar] [PubMed]
- Hood S, Amir S. The aging clock: circadian rhythms and later life. J Clin Investig. 2017;127:437-46. [CrossRef] [Google scholar] [PubMed]
- Miner B, Kryger MH. Sleep in the Aging Population. Sleep Med Clin. 2017;12(1):31-38. doi:10.1016/j.jsmc.2016.10.008. PMID: 28159095 [CrossRef] [Google scholar] [PubMed]
- Yoon IY, Kripke DF, Elliott JA, Youngstedt SD, Rex KM, Hauger RL. Age-related changes of circadian rhythms and sleep-wake cycles. J Am Geriatr Soc. 2003;51:1085-91. [CrossRef] [Google scholar] [PubMed]
- Blackwell T, Yaffe K, Laffan A, Ancoli-Israel S, Redline S, Ensrud KE, et al. Associations of objectively and subjectively measured sleep quality with subsequent cognitive decline in older community-dwelling men: The MrOS sleep study. Sleep. 2014;37:655-63. [CrossRef] [Google scholar] [PubMed]
- Mander BA, Winer JR, Jagust WJ, Walker MP. Sleep: a novel mechanistic pathway, biomarker, and treatment target in the pathology of Alzheimer’s Disease? TINS. 2016;39:552-66. [CrossRef] [Google scholar] [PubMed]
- De Boer A, Horst GJT, Lorist MM. Physiological and psychosocial age-related changes associated with reduced food intake in older persons. Ageing Res Rev. 2013;12:316-28. [CrossRef] [Google scholar] [PubMed]
- Landi F, Calvani R, Tosato M, Martone AM, Ortolani E, Savera G, et al. Anorexia of aging: risk factors, consequences, and potential treatments. Nutrients. 2016;8:69-79. [CrossRef] [Google scholar] [PubMed]
- Shilsky J, Bloom DE, Beaudreault AR, Tucker KL, Keller HH, Freund-Levi Y, et al. Nutritional considerations for healthy aging and reduction in age-related chronic diseases. Adv Nutr. 2017;8:17-26. [CrossRef] [Google scholar] [PubMed]
- Martone AM, Onder G, Vetrano DL, Ortolani E, Tosato M, Marzettie E, et al. Anorexia of aging: a modifiable risk factor for frailty. Nutrients. 2013;5:4126-33. [CrossRef] [Google scholar] [PubMed]
- Azhar G, Wei JY. Nutrition and cardiac cachexia. Curr Opin Clin Nutrition Metabol Care. 2006;9:18-23. [CrossRef] [Google scholar] [PubMed]
- Azhar G, Wei JY. The demographics of aging and its impact on the cardiovascular health. Curr Cardiovascular Risk Reports. 2015;4:1-6. [CrossRef] [Google scholar]
- Cesari M, Vellas B, Hsu F-C, Newman AB, Doss H, King C, et al. A physical activity intervention to treat the frailty syndrome in older persons—results from the LIFE-P study. J Gerontol Series A: Biol Sci Med Sci. 2015;70:216-22. [CrossRef] [Google scholar] [PubMed]
- Conroy S, Elliott A. The frailty syndrome. Medicine. 2017;45:15-18. [CrossRef] [Google scholar]
- Lopez-Otin C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153:1194-217. [CrossRef] [Google scholar] [PubMed]
- Newgard CB, Pessin JE. Recent progress in metabolic signalling pathways regulating aging and life span. J Gerontol A Biol Sci Med Sci. 2014;69:S21-7. [CrossRef] [Google scholar] [PubMed]
- Barbado EL, Aquilano K. Feast and famine: adipose tissue adaptations for healthy aging. Ageing Res Rev. 2016;28:85-93. [CrossRef] [Google scholar] [PubMed]
- Calcada D, Vianello D, Giampieri E, Sala C, Castellani G, de Graaf A, et al. The role of low-grade inflammation and metabolic flexibility in aging and nutritional modulation thereof: a systems biology approach. Mech Ageing Devel. 2014;136-7:138-147. [CrossRef] [Google scholar] [PubMed]
- Franceschi C, Capri M, Moni D, Giunta S, Olivieri F, Sevini F, et al. Inflammaging and anti-inflammaging: a systemic perspective on aging and longevity emerged from studies in humans. Mech Ageing Dev. 2007;128:92-103. [CrossRef] [Google scholar] [PubMed]
- Ingliosi AM, Opp MR, Krueger JM. Sleep and immune function: glial contributions and consequences of aging. Curr Opin Neurobiol. 2013;23:806-11. [CrossRef] [Google scholar] [PubMed]
- Nedeltcheva AV, Scheer FAJL. Metabolic effects of sleep disruption, links to obesity and diabetes. Curr Opin Endocrinol Diabetes Obes. 2014;21:293-8. [CrossRef] [Google scholar] [PubMed]
- Potter GCM, Skene DJ, Arendt J, Cade JE, Grant PJ, Hardie LJ. Circadian rhythm and sleep disruption: causes, metabolic consequences, and counter measures. Endocrine Rev. 2016;37:584-608. [CrossRef] [Google scholar] [PubMed]
- Zmora N, Bashiardes S, Levy M, Elinav E. The role of the immune system in metabolic health and disease. Cell Metabolism. 2017;25:506-21. [CrossRef] [Google scholar] [PubMed]
- Gruver AL, Hudson LL, Sempowski GD. Immunosenescence of ageing. J Pathol. 2007;211(2):144-56. PMID: 17200946 [CrossRef] [Google scholar] [PubMed]
- Prinz PN. Age impairments in sleep, metabolic and immune functions. Exp Gerontol. 2004;39:1739-43. [CrossRef] [Google scholar] [PubMed]
- Grice EA, Segre JA. The human microbiome: our second genome. Annu Rev Genomic Hum Genet. 2012;13:151-70. [CrossRef] [Google scholar] [PubMed]
- Sankar SA, Lagier JC, Pontarotti P, Raoult D, Fournier PE. The human gut microbiome, a taxonomic conundrum. Syst Appl Microbiol. 2015;38(4):276-86. PMID: 25864640 [CrossRef] [Google scholar] [PubMed]
- Kumar M, Babaei P, Ji B, Nielsen J. Human gut microbiota and healthy aging: Recent developments and future prospective. Nutr Healthy Aging. 2016;4(1):3-16. PMID: 28035338 [CrossRef] [Google scholar] [PubMed]
- De Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA. 2010;107: 14691-6. [CrossRef] [Google scholar] [PubMed]
- Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, et al. Human gut microbiome viewed across age and geography. Nature. 2012;486:222-7. [CrossRef] [Google scholar] [PubMed]
- Palmer C, Bik EM, DiGiulio DB, Relman DA, Brown PO. Development of the human infant intestinal microbiota. PLoS Biol. 2007;5(7):e177. PMID: 17594176 [CrossRef] [Google scholar] [PubMed]
- Lynch SV, Pedersen O. The human intestinal microbiome in health and disease. N Engl J Med. 2016;375:2369-79. [CrossRef] [Google scholar] [PubMed]
- Johnson EL, Heaver SL, Walters WA, Ley RE. Microbiome and metabolic disease: revisiting the bacterial phylum Bacteroidetes. J Mol Med (Berl). 2016;95(1):1-8. PMID: 27900395 [CrossRef] [Google scholar] [PubMed]
- Bik EM. The hoops, hopes and hypes of human microbiome research. Yale J Biol Med. 2016;89:363-73. [Google scholar]
- Wang Y, Kasper LH. The role of microbiome in central nervous system disorders. Brain, Behav Immunity. 2014;38:1-12. [CrossRef] [Google scholar] [PubMed]
- Luna RA, Foster JA. Gut brain axis: diet microbiota interactions and implications for modulation of anxiety and depression. Curr Opin Biotechnol. 2015;32:35-41. [CrossRef] [Google scholar] [PubMed]
- Reynolds AC, Paterson JL, Ferguson SA, Stanley D, Wright KP, Dawson D. The shift work and health research agenda: considering changes in gut microbiota as a pathway linking shift work, sleep loss and circadian misalignment, and metabolic disease. Sleep Med Rev. 2016; doi:10.1016/j.smrv.2016.06.009. [CrossRef] [Google scholar] [PubMed]
- Rea K, Dinan TG, Cryan JF. The microbiome: a key regulator of stress and neuroinflammation. Neurobiol of Stress. 2016;4:23-33. [CrossRef] [Google scholar] [PubMed]
- Bressa C, Bailen-Andrino M, Perez-Santiago J, Gonzalez-Soltero, R, Perez M, Montalvo-Lominchar MG, et al. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE. 2017;12:e0171352. [CrossRef] [Google scholar] [PubMed]
- Arboleya S, Watkins C, Stanton C, Ross RP. Gut Bifidobacteria populations in human health and aging. Front Microbiol. 2016;7:1204. [CrossRef] [Google scholar] [PubMed]
- Vaiserman AM, Koliada AK, Marotta F. Gut microbiota: a player in aging and a target for anti-aging intervention. Ageing Res Rev. 2017;35:36-45. [CrossRef] [Google scholar] [PubMed]
- Jackson MA, Jeffery IB, Beaumont M, Bell JT, Clark AG, Ley RE et al. Signatures of early frailty in the gut microbiota. Genome Med. 2016;8:8-19. [CrossRef] [Google scholar] [PubMed]
- Sampson TR, Mazmanian SK. Control of brain development, function, and behaviour by the microbiome. Cell Host & Microbe. 2015;17:565-76. [CrossRef] [Google scholar] [PubMed]
- Thaiss CA, Zmora N, Levy M, Elinav E. the microbiome and innate immunity. Nature. 2016;535:65-74. [CrossRef] [Google scholar] [PubMed]
- O’Mahony SM, Clarke G, Borre YE, Dinan TG, Cryan JF. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav Br Res. 2015;277:32-48. [CrossRef] [Google scholar] [PubMed]
- Sandhu KV, Sherwin E, Schellekens H, Stanton C, Dinan TG, Cryan JF. Feeding the microbiota-gut-brain axis: diet, microbiome, and neuropsychiatry. Transl Res. 2017;179:223-44. [CrossRef] [Google scholar] [PubMed]
- Carabotti M, Scirocco A, Maselli MA, Severi C. the gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Ann Gastroenterol. 2015;28:203-9. [Google scholar]
- Dinan TG, Cryan JF. Gut instincts: microbiota as a key regulator of brain development, aging and neurodegeneration. J Physiol. 2017;595:489-503. [CrossRef] [Google scholar] [PubMed]
- Tremlett H, Bauer KC, Appel-Cresswell S, Finlay BB, Waubant E. The gut microbiome in human neurological disease: a review. Ann Neurol. 2017; doi:10.1002/ana.24901. [CrossRef] [Google scholar] [PubMed]
- Grenham S, Clarke G, Cryan JF, Dinan TG. Brain-gut-microbe communication in health and disease. Front Physiol. 2011;2:94. [CrossRef] [Google scholar] [PubMed]
- Kennedy PJ, Cryan JF, Dinan TG, Clarke G. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology. 2017;112(Pt B):399-412. [Google scholar]
- Bauer KC, Huus KE, Finlay BB. Microbes and the mind: emerging hallmarks of the gut microbiota-brain axis. Cell Microbiol. 2016;18:632-44. 86. Powell N, Walker MM, Talley NJ. The mucosal im [CrossRef] [Google scholar] [PubMed]
- Powell N, Walker MM, Talley NJ. The mucosal immune system: master regulator of bidirectional gut-brain communications. Nat Rev Gastroenerol & Hepatol. 2016;14:143-59. [CrossRef] [Google scholar] [PubMed]
- Mittal R, Debs LH, Patel AP, Nguyen D, Patel K, O’Connor G et al. Neurotransmitters: the critical modulators regulating gut-brain axis. J Cell Physiol. 2016; doi:10.1002/jcp.25518. [CrossRef] [Google scholar] [PubMed]
- Million M, Larauche M. Stress, sex and the enteric nervous system. Neurogastroenterol Motility. 2016;28:1283-9. [CrossRef] [Google scholar] [PubMed]
- Hyland NP, Cryan JF. A Gut Feeling about GABA: Focus on GABA(B) Receptors. Front Pharmacol. 2010;1:124. PMID: 21833169 [CrossRef] [Google scholar] [PubMed]
- De Palma G, Collins SM, Bercik P. The microbiota-gut-brain axis in functional gastrointestinal disorders. Gut Microbes. 2014;5:419-29. [CrossRef] [Google scholar] [PubMed]
- McCann H, Cartwright H, Halliday GM. Neuropathology of -synuclein propagation and Braak hypothesis. Movement Dis. 2016;31:152-60. [CrossRef] [Google scholar] [PubMed]
- Jeffery IB, Lynch DB, O'Toole PW. Composition and temporal stability of the gut microbiota in older persons. ISME J. 2016;10:170-82. [CrossRef] [Google scholar] [PubMed]
- Ljungberg B, Nilsson-Ehle I, Edlund C, Nord CE. Influence of ciprofloxacin on the colonic microflora in young and elderly volunteers: no impact of the altered drug absorption. Scand J Infect Dis. 1990;22(2):205-8. [CrossRef] [Google scholar] [PubMed]
- Hopkins MJ, Sharp R, Macfarlane GT. Age and disease related changes in intestinal bacterial populations assessed by cell culture, 16S rRNA abundance, and community cellular fatty acid profiles. Gut. 2001 Feb;48(2):198-205. [CrossRef] [Google scholar] [PubMed]
- Bartosch S, Fite A, Macfarlane GT, McMurdo ME. Characterization of bacterial communities in feces from healthy elderly volunteers and hospitalized elderly patients by using real-time PCR and effects of antibiotic treatment on the fecal microbiota. Appl Environ Microbiol. 2004;70(6):3575-81. PMID: 15184159 [CrossRef] [Google scholar] [PubMed]
- Bindels LB, Delzenne NM. Muscle wasting: the gut microbiota as a new therapeutic target? International J Biochem Cell Biol. 2014;45:2186-90. [CrossRef] [Google scholar] [PubMed]
- Nagpal R, Yadav H, Marotta F. Gut microbiota: the next-gen frontier in preventive and therapeutic medicine? Front Med. 2014;1:15. [CrossRef] [Google scholar] [PubMed]
- Duffy LC, Raiten DJ, Hubbard VS, Starke-Reed P. Progress and challenges in developing metabolic footprints from diet in human gut microbial cometabolism. J Nutr. 2015;145:1123S-30S. [CrossRef] [Google scholar] [PubMed]
- Broussard JL, Van Cauter E. Disturbances of sleep and circadian rhythms: novel risk factors for obesity. Curr Opin Endocrinol Diabetes Obes. 2016;23:353-9. [CrossRef] [Google scholar] [PubMed]
- Voigt RM, Forsyth CB, Green SJ, Mutlu E, Engen P, Vitaterna MH, et al. Circadian disorganization alters intestinal microbiota. PLoS One. 2014;9:e97500. [CrossRef] [Google scholar] [PubMed]
- Mukherji A, Kobiita A, Ye T, Chambon P. Homeostasis in intestinal epithelium is orchestrated by the circadian clock and microbiota cues transduced by TLRs. Cell. 2013;153:812-27. [CrossRef] [Google scholar] [PubMed]
- Salles N. Basic mechanisms of the aging gastrointestinal tract. Dig Dis. 2007;25:112-7. [CrossRef] [Google scholar] [PubMed]

