

Molecular Cytogenetic Characterization of Two Murine Colorectal Cancer Cell Lines
Hans Rhode 1![]() , Thomas Liehr 1,*
, Thomas Liehr 1,*![]() , Nadezda Kosyakova 1
, Nadezda Kosyakova 1![]() , Martina Rinčić 2
, Martina Rinčić 2![]() , Shaymaa S. Hussein Azawi 1
, Shaymaa S. Hussein Azawi 1![]()
- Jena University Hospital, Friedrich Schiller University, Institute of Human Genetics, Jena, Germany
- Department for Functional Genomics, Centre for Translational and Clinical Research, University Hospital Centre Zagreb, University of Zagreb School of Medicine, Zagreb, Croatia
* Correspondence: Thomas Liehr![]()
Received: July 30, 2018 | Accepted: September 12, 2018 | Published: September 20, 2018
OBM Genetics 2018, Volume 2, Issue 3 doi:10.21926/obm.genet.1803037
Academic Editors: Joep Geraedts
Special Issue: Applications of Fluorescence in Situ Hybridization
Recommended citation: Rhode H,Liehr T,Kosyakova N,Rinčić M,Azawi SSH. Molecular Cytogenetic Characterization of Two Murine Colorectal Cancer Cell Lines. OBM Genetics 2018;2(3):037; doi:10.21926/obm.genet.1803037.
© 2018 by the authors. This is an open access article distributed under the conditions of the Creative Commons by Attribution License, which permits unrestricted use, distribution, and reproduction in any medium or format, provided the original work is correctly cited.
Abstract
(1) Background: Colorectal cancer (CRC) is the third most common cancer in human and the fourth leading cause of adult man’s death. Murine tumor cell lines have been established as a model system for CRC, but their cytogenetic properties have so far been only poorly understood. (2) Methods: The two murine colon tumor cell lines CMT-93 and CT26 (also called CT26.WT, CT-26 or CT-26 WT) were investigated in this study by molecular cytogenetic methods, i.e. by multicolor-fluorescence in situ hybridization (mFISH), murine multicolor banding (mcb), and array-based comparative genomic hybridization (aCGH). The thereby characterized chromosomal imbalances and chromosomal breakpoints were compared with those of human CRC by in-silico-translation of murine data into the human genome. (3) Results: CMT-93 and CT26 showed a hyperdiploid and hypertriploid karyotype, respectively. While only clonal aberrations of chromosomes 2, 5, 8, and X were observed for CMT-93, there was greater variability of chromosomal imbalances observed in CT26. Both cell lines tended to form dicentric and neocentric chromosomes and showed 17 (CMT-93) and 28 tumor-associated breakpoints (CT26). Interestingly, imbalances found were almost exclusively gains in somatic chromosomes. Also Y-chromosome was lost in CMT-93 as was loss of one of the X-chromosomes in CT26. In-silico-translation of the in both cell lines observed chromosomal imbalances showed a high agreement with the most frequently in human CRC observed metastatic amplifications. (4) Conclusions: This study revealed that murine tumor cell lines CMT-93 and CT26 are models for human CRCs of advanced tumor stages. This information is important to apply CMT-93 and CT26 for the correspondingly suited questions to be studied.
Keywords
Colorectal cancer; CRC; CMT-93; CT26; murine tumor; cell lines; multicolor-fluorescence in situ hybridization; mFISH; murine multicolor banding; mcb; array-based comparative genomic hybridization; aCGH
1. Introduction
Colorectal cancer (CRC) is the third most common human cancer worldwide and the fourth most common cause of cancer-related deaths in adults [1]. Mutations in specific genes, like oncogenes, tumor suppressor genes, and genes related to DNA-repair mechanisms, can lead to the onset of CRC [2]. In addition, other genetic changes can result in a loss of genomic stability [3]. These “other” changes are now being considered as a factor possibly more important, especially in relation to CRC [2], than gene mutations [4].
Most commonly, CRC is localized in the rectum or the sigmoid colon. According to the classification of the Union International Contre Le Cancer (UICC), the four stages of CRC are distinguished as follows: (I) limitation on the intestinal wall; (II) growth beyond the intestinal wall; (III) lymphatic metastasis; and (IV) distant metastases, mainly in liver or lung [5]. With the elucidation of the molecular pathways involved in tumorigenesis, novel models of human-cancer development have been created to further advance our understanding of CRC, increase our ability to evaluate potential therapies, and positively affect patients’ outcomes [1,6].
In humans, it is not possible to fully study primary tumors, distant metastatic sites (one-at-a-time and in one organism), or to carry out genetic manipulation (e.g., the spatial and temporal expression levels of certain genes). To overcome this barrier, animal models have been established and have been shown to be a powerful tool in cancer research. The mouse provides an excellent platform for modelling cancer in the mammalian system. Due to its comparatively easy breeding (small body size, short regeneration time) and its basic similarity to humans in terms of genetics and physique, it offers some advantages over other animal models [7]. Thus, murine models have been extensively used in cancer studies (e.g. to directly monitor the metastatic progression in CRCs) [1]. In addition to the mouse as a whole organism and the development of embryonic stem cell lines, murine tumor cell lines were introduced into research in the 1970s [7,8]. As stated previously [9,10,11], even though murine tumor cell lines have been used in many experiments, including those for drug development [12], murine tumor cells are, overall, not well characterized genetically.
Despite the fact that CMT-93 and CT26 are immortal, easy to cultivate, and readily available from repositories, and were induced in 1975 and 1978 by use of alkylating agents, a gap in knowledge exists. They are most often used in studies involving carcinogenesis and chemotherapeutics [6,12,13]. Beginning in 1983 and continuing in 2018, CMT-93 was used in 72 studies listed in PubMed (https://www.ncbi.nlm.nih.gov/pubmed/?term=CMT-93), and CT26 (also called CT26.WT, CT-26 or CT-26 WT) was used in approximately 1,200 studies listed in PubMed (https://www.ncbi.nlm.nih.gov/pubmed/?term=CT26; https://www.ncbi.nlm.nih.gov/pubmed/?term=CT26.WT). Even though their pathobiologic characteristics are very similar to those of human CRCs [3], we must once again stress that most of these studies were done without any available genetic information on the two cell lines. Recently, one study involving CMT-93 was conducted, analyzing its expression at the cDNA level, and another study was published characterizing CT26 by next-generation sequencing of [1,14].
To close this gap in knowledge, we did a comprehensive (molecular) cytogenetic description and in silico- (= genome browser based-) translation to the human genome for both cell lines. Thus, for the first time, it is possible to characterize the subtype of human CRCs, CMT-93, and CT26, that can be used for as models.
2. Materials and Methods
2.1. Murine CRC Cell Lines
The fibroblast-like growing, epithelial, colon-derived cell lines CMT-93 and CT26 were obtained from American Type Culture Collection (ATCC® CCL-223 and CRL-2638, respectively). These cells, originally derived from colon, grow adherently in RPMI-1640 medium with 2mM L-glutamine, 10mM HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid), and 10% fetal-calf serum in the presence of antibiotics. For this study, we split the cells into two portions. Flask 1 was worked up cytogenetically, i.e., cells were subjected to colcemid at an end-concentration of 0.1µg/ml for 2-3 hours. Subsequently the cells were harvested by standard trypsin treatment, put into a 15ml tube, and hypotonically treated in 0.075M KCl for 20 minutes. Next, they were fixed, and cytogenetic work-up was completed. Overall metaphase spreads were prepared according to standard procedures[15]. Cells from Flask 2 were used to extract whole-genomic DNA by Blood & Cell Culture DNA Midi Kit (Qiagen, Düsseldorf, Germany) according to manufacturer’s instructions.
According to written statements from the ethical committee (medical faculty) and the Animal Experimentation Commission of the Friedrich Schiller University, there are no ethical statements necessary for cases involving the use of murine tumor-cell lines, like CMT-93 and CT26.
2.2. Molecular Cytogenetics
Fluorescence in situ hybridization (FISH) was performed as previously described [10]. “SkyPaintTM DNA Kit M-10 for Mouse Chromosomes” (Applied Spectral Imaging, Edingen-Neckarhausen, Germany) was applied for multicolor-FISH (mFISH) with whole chromosome paints, and murine chromosome-specific multicolor banding (mcb) probe mixes for FISH-banding [16]. At least 30 metaphases were documented and analyzed for each probe set on a Zeiss Axioplan microscope, equipped with ISIS software (MetaSystems, Altlussheim, Germany). Array-based comparative genomic hybridization (aCGH) was completed according to standard procedures with “SurePrint G3 Mouse CGH Microarray, 4x180K” (Agilent Technologies) [10].
2.3. Data Analysis
The regions of imbalances and breakpoints in CMT-93 and CT26 were characterized after analyses of aCGH and mcb data, and aligned with their human homologous regions using Ensembl and the UCSC Genome Browser, as previously described [9]. To align the data we obtained with the human CRCs, data was collected from Diep et al. [17], Golovko et al. [12], Gordon et al. [5], Knutsen et al. [13], Kloosterman et al. [22], Marmol et al. [2], and Skuja et al. [18].
3. Results
Seventeen tumor-associated breakpoints were detected in CMT-93, and twenty-eight in CT26. Both cell lines showed dicentric (dic) and “neocentric” (neo) chromosomes. For the latter, it must be admitted that either these are really neocentric chromosomes, or they are derivative chromosomes with an “original centromeric sequence” derived from band A1, which, for technical reasons, could not be attributed to the specific chromosome. If the latter is valid, then all derivatives denominated as ‘neo’ would have an interstitial deletion from sub-band A1 up to the first band present of this specific chromosome. Both scenarios, formation of a neocentric chromosome or formation of a complex rearranged derivative chromosome with centromere-near interstitial deletion, are unique and not yet reported in murine cell lines.
3.1. CMT-93
CMT-93 can be considered as a hyperdiploid cell line with a tendency for polyploidization (6-20% of the cells). Besides, CMT-93 has structural aberrations in chromosomes 2, 5, 8, and X (Figure 1A); two different derivatives of chromosome 8; with two different breakpoints, 8B1 and 8C2 (Figure 1B). The cells of CMT-93 could attribute into two main clones, each of which is present in about 50% of the cells. Examples of ‘neocentric’ derivatives are shown in Figure 2.
Karyotype of clone 1.
Tex48~56,XXY, +der(2)(neo::G2→qter), +3, +der(4)(neo::C6→C7:), +5, +6, +der(8)(pter→D1::D1→C2:), +der(8)(neo::B1.1→qter), +der(8)(neo::C2→qter)x2, +11, +12, +13,dic(14;15)(A1;A1), +15, +17, +18, +19, +19. Two metaphases (= 13% of clone 1) showed a tetraploid karyotype.
Karyotype of clone 2.
49~54,XXY, +der(2)(neo:G2→qter), +3, +der(4)(neo::C6→C7:), +5, +6, +der(8)(pter→D1::D1→C2:), +der(8)t(2;8)(B1.1;E1), +der(8)(neo:C2→qter), +11, +13,dic(14;15)(A1;A1), +15, +17, +19.
Due to ongoing karyotype evolution in clone 2 two small subclones were detected:
- in about 4% of the cells with main karyotype of clone 2 instead of a chromosome 2 and 5 there are a der(2)t(2;5)(C3;E3), and a del(5)(E3), and
- in about 12% 4% of the cells with main karyotype of clone 2 one normal X-chromsome was replaced by a der(X)(pter→F5::A3→qter); also an additional chromosome 12 is present.

Figure 1 (A) Murine multicolor banding (mcb) was applied on chromosomes of the CMT-93 cell line. Typical pseudocolor banding is for all 21 different murine chromosomes, apart from the Y-chromosome, are shown for clone 2. This figure depicts the summary of 20 chromosome-specific FISH-experiments, overall. Two derivative chromosomes consist of two different chromosomes, each. These are highlighted by frames and shown twice in this summarizing karyogram; (B) There are two different derivatives of normal chromosome 8 (#8) in CMT-93. Either they are neocentrics, which acquired new centromeres in B1.1 or C2, or they are due to interstitial deletions; the latter option is given in the corresponding schemes of the derivative chromosomes.
3.2. CT26
CT26 is a triploid cell line with a tendency for tetraploidization (Figure 2). Genomic instability expressed as changes in chromosome numbers is clearly more expressed than in CMT-93, which made the identification of a main clone more difficult. Even though mFISH, mcb, and aCGH were applied, there remained unidentifiable marker chromosomes in these karyotypes. Most likely, they are derivatives from non-characterizable, heteochromatic bands A1 from any of the 21 murine chromosomes.
Karyotype of mainclone.
59~61,XX,dic(1;14)(A1;A1), +1, +der(2)t(2;9)(neo::2H2→2C3::9E2→9qter),der(3)t(3;15)(3pter→3H4::neo::3F3→3qH4::3H4→3E1::15A1→15qter),dic(5;15)(A1;A1),dic(5;19)(A1;A1), +dic(5),dic(6;6)(A1;A1), +der(6)(3qter→3A1::6A3→6A1:6A1→6G3::10C3→10qter), +der(7)t(7;19)(D1;D1), +8,der(9)t(2;9)(H2;E1),dic(12;12)(A1;A1),dic(13;13)(A1;A1),dic(13;17)(A1;A1), der(13)(neo:A3→C3:),dic(14;14)(A1;A1), +17, +19, +mar1x2.
This clone was found in about 12% of the cells, and another 26% additionally had: +der(18)(pter→C::B3→qter), +19, +mar2x2, +mar3x3.
In addition, there were 20% of cells with the karyotype of the main clone and an additional dic(X;12)(A5;A1). However, the latter dicentric is further changed to dic(X;12)(A1;A1) in the remainder 42% of the cells.
Still, especially in the 42% of the CT26 cells, there were additionally small subclones (5-10%), with e.g. dic (1;14)(A1;A1), dic(5;15)(A1;A1), der(3)t(3;15), or der(18)(pter→C::C→qter). Also there is evidence from mFISH analyses that der(6)(3qter→3A1::6A3→6A1:6A1→6G3::10C3→10qter) developed from a der(6)t(6;10) only detectable in mFISH and not in mcb analyses. Tetraploid karyotypes were found in 20-30% of CT26 cells.
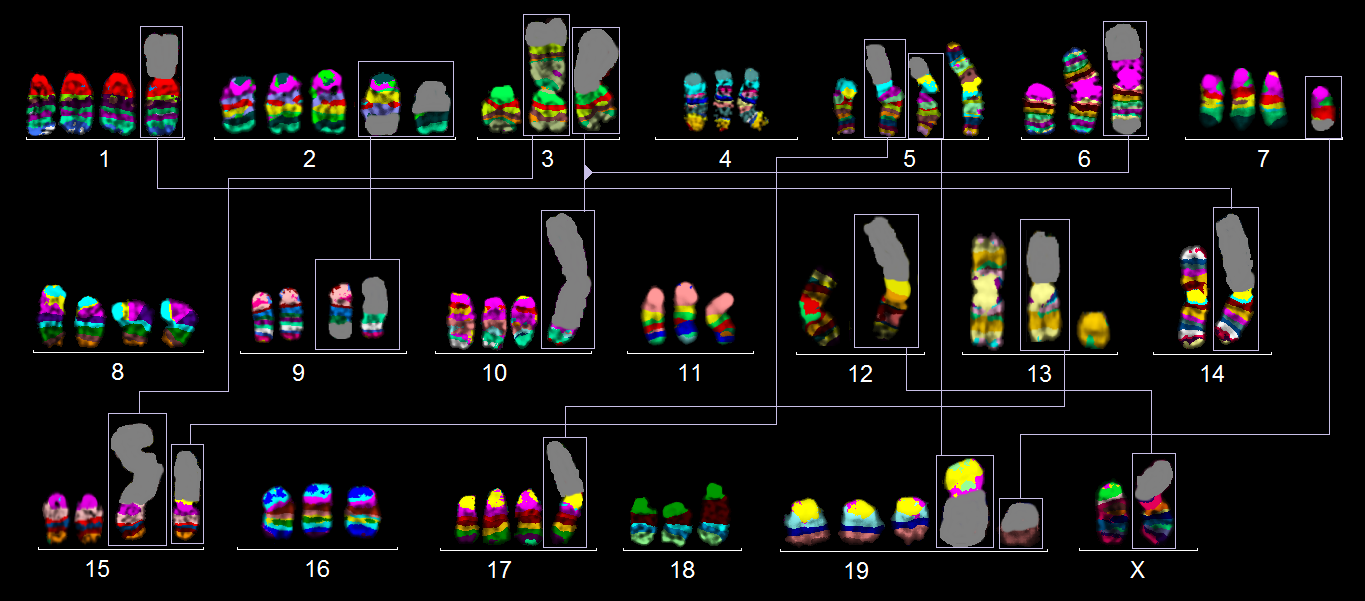
Figure 2 results of murine multicolor banding (mcb) for cell-line CT26, as shown in Figure 1A for CMT-93. Here, 10 derivative chromosomes consist of two different chromosomes, each being highlighted by frames and shown twice in this summarizing karyogram.
3.3. aCGH and In-silico-translation
The results of the aCGH are detailed in Table 1a for CMT93 and Table 1b for CT26. Overall, these results were consistent with the results of molecular cytogenetics for both cell lines. FISH- and aCGH-results are summarized in Figure 3. For this study, only imbalances larger than 3.5 megabase pairs were included in the evaluation. Results of translation of the aberrations found in both murine cell lines and the human genome are shown graphically in Figure 4.
Table 1 The regions of gain and loss of copy numbers, as well of breakpoints of balanced rearrangements, observed in CMT-93 and CT26 and the corresponding homologue regions in humans, are listed as cytoband and position (GRCh37/hg19).
|
Table 1a: CMT-93 |
|||
|
region |
gain |
homologue region in human |
|
|
cytoband |
position (GRCh37/hg19) |
||
|
2qE1-qG2 |
+1 |
20p11.1-p13 11p11.2-p14.2 15q13.3-q21.2 2q13 2q13 2q11.11–q11.2 |
20:1736101-25606620 11:26296397-48658712 15:32906987-51298173 2:112973390-113650007 2:111483204-112960231 2:95642277-97040617 |
|
2qG2-qH4 |
+2 |
20q13.32-q13.33 20p11.21-p13.32 20p13 |
20:58148222-62907435 20:29933153-58056214 20:102147-1447942 |
|
3qA1-qH4 |
+1 |
7q11.23 8q21.11-q21.3 8q12.3-q13.1 3q24-q25.1 3q26.2-q26.32 3q26.32-q27.1 4q27-q31.1 21p11.2 9q13 9p11.2 9p12 13q13.2-q14.11 3q25.1-q26.2 4q31.3-q32.2 4q26-q27 4q22.3-q26 1q21.1-q23.1 1p11.2-p22.1 1p22.2-p31.3 |
7:76282730-76575579 8:76197937-87035414 8:64075897-67315825 3:148467535-148965303 3:167857105-178105807 3:178137517-182818465 4:122242382-141190230 21:10369840-10592667 9:68139917-68294733 9:45446787-46098069 9:42028291-42246766 13:34463185-41254213 3:149055816-167822106 4:150966383-163096512 4:119596924-120703320 4:95284699-119338945 1:144676687-158154741 1:93905157-120696915 1:68589539-89738135 |
|
4qC6-qC7 |
+1 |
1p32.1-p36.33 |
1:894315-59012766 |
|
5qA1-qG3 |
+1 |
1p22.1-p22.2 13q12.13-q13.2 2q23.2-q23.3 2q23.3 2q23.3 4q12-q22.1 4p16.3-p11.1 4p16.3 4p16.3 12q24.33 12q24.31-q24.33 12q24.11-q24.31 12q23.3-q24.11 18p11.32 22q11.23-q12.1 22q12.2-q12.3 7p22.1-p22.3 7p15.3 7q11.23-q21.2 7q22.1-q22.3 7q36.1-q36.3 7p15.3 7q36.1 7q21.3-q22.1 7q22.1 7q11.23 7q11.21-q11.23 7p11.2 |
1:89950168-93744300 13:26784894-34260463 2:27759882-29024462 2:27256674-27749458 2:26394466-27256616 4:52689038-89000187 4:4184743-49083612 4:1109142-3830658 4:493106-1023731 12:132378991-133522542 12:121577100-132336561 12:110488793-121497537 12:108325357-110486420 18:683166-844532 22:25201765-29156283 22:32022117-32511666 7:169204-6771649 7:22903126-23240676 7:76702246-92676514 7:102331617-105210094 7:150588373-157179154 7:22595306-22772664 7:149628652-149959756 7:97598308-99229367 7:99552841-102191754 7:72536306-76149827 7:66808098-72045725 7:56019352-56184138 |
|
6qA1-qG3 |
+1 |
1p31.3 22q11.1-q11.21 4q27 4q22.1-q22.3 10q11.21-q11.22 2p11.2-p13.3 2p11.2 3p25.2-pter 3p25.1-p25.2 3p12.3-p14.1 3q21.3 3q22.1 7p14.3-p15.3 7q31.1-q36.1 7p21.3-p22.1 7p21.3 7p11.2 12p11.21-p13.31 12p13.31-p13.33 12p13.31 12p13.33 |
1:67631910-68317098 22:17565811-18659740 4:121018693-122194687 4:89178698-95273100 10:43277986-46218167 2:68715037-87095119 2:88302422-89174373 3:61304-12897767 3:12939278-15163105 3:64017713-75322601 3:125725101-129038484 3:129094932-129632650 7:23254035-33103246 7:112138919-149583263 7:7132996-12536829 7:92745197-97502117 12:31424829-32537434 12:9901365-30943693 12:2903120-7695890 12:8071763-9214464 12:66113-2823666 |
|
8qA1-qC2 |
+1 |
4q32.2-qter 19p13.11-p13.12 19p13.2 13q33.1-qter 13q14.3 8p22-p23.1 8p23.1 8p11.23-p12 8p11.21-p11.23 8p23.1-p23.2 8p23.2-p23.3 |
4:163504024-190884657 19:16163040-19774937 19:7112183-8071013 13:103533915-115092930 13:52435459-53211718 8:12579073-17958954 8:8108776-9640417 8:29190466-36677574 8:36716542-42505949 8:5368147-6693649 8:591286-5358752 |
|
8qC2-qD3 |
+3 |
4q31.1-q31.23 16q12.1-q22.1 19p13.12-p13.2 |
4:141251922-150892329 16:46693273-69976105 19:12745060-14683008 |
|
8D3-qter |
+1 |
16q22.1-qter 10p11.21-p11.22 1q42.13-q32.3 |
16:70109527-90110030 10:33112469-35152269 1:229404294-235324774 |
|
11qA1–qE2 |
+1 |
22q12.1-q12.2 7p11.2-p13 2p13.3-p16.2 5q35.2 5 q33.2-q35.1 5q35.3 5q23.3-q31.1 5 q33.1-q33.2 1q42.13 17p11.2 17p12-p13.3 17q11.1-q11.2 17p11.2-p12 17q21.32-q23.2 17q12-q21.31 17q21.31- q21.32 17q23.2-q24.1 17q21.32 17q24.1-q24.2 17q24.2-qter 17q11.2-q12 |
22:29251511-32022116 7:43906144-55317931 2:53882943-68694726 5:172736725-173663599 5:154331837-171932313 5:177531363-180585244 5:130484032-134063627 5:150381711-154330989 1:227919753-228703212 17:16917258-21343117 17:2-15625804 17:25525650-28853901 17:29058377-36200511 17:45560334-60326198 17:36351926-43638822 17:43706746-45150591 17:60483588-62760387 17:45188646-45518436 17:62990972-66110690 17:66224207-81175056 17:15731601-16472951 |
|
12qA1.1-qF2 |
+1 |
2p23.3-p25.1 2p25.1 2p25.1 2p25.1-pter 7q22.3-q31.1 7p21.1-p21.3 7q31.1 14q12-q22.1 14q23.1-32.33 7q36.3 7p15.3-21.1 |
2:10303009-26361943 2:9354723-9994801 2:9996101-10284917 2:140908-9278318 7:105210238-107772185 7:12561752-19748810 7:107772206-112136146 14:25157192-52251174 14:58666612-106375879 7:157225645-158937901 7:19761201-22528893 |
|
15qA1-qF3 |
+1 |
5p12-p15.31 8q22.1-q24.3 22q12.3-qter 12q12-q13.2 12p11.1 |
5:8927745-42888975 8:97446632-146158346 22:35962951-51222438 12:38607141-55072925 12:33476533-34210697 |
|
17qA1-qE5 |
+1 |
18p11.32 18p11.22-p11.32 2p16.2-p16.3 2p16.3-p23.2 2q12.2-q12.3 3p24.3-p25.1 5q21.2-q22.1 19p13.3 6p12.3-p21.2 6 p21.32-p22.1 6p21.2-p21.31 6 q27 6 q25.3-q27 6q25.2-q25.3 5q15-q21.1 16p13.3 5q35.1 21q22.3 19p13.12 19p13.2 |
18:861722-2534400 18:2534401-9972541 2:51709987-53282184 2:29033520-51699597 2:107383985-108798215 3:16307846-20231899 5:102759315-110063021 19:4229082-6862967 6:39266498-49681826 6:29322703-33297218 6:33359177-39058058 6:167859539-170893754 6:160103032-166797236 6:155053083-160101646 5:96202316-98405239 16:222880-3208490 5:171946752-172722349 21:43490502-45122943 19:15270296-15808207 19:8366687-8811037 |
|
19qA-qD3 |
+2 |
11q12.1-q13.3 9q21.11-q21.31 9p24.1-pter 2q13 10q11.23-q21.1 10q23.2-q26.11 |
11:57844834-68709722 9:69086307-82777364 9:51374-6659223 2:114171139-114321953 10:51917603-54540082 10:89234113-121219507 |
|
XqA4-qF5 |
+1 |
Xp22.11-p22.2 Xp11.21-p11.22 Xp22.2 Xq11.1-q23 Xp21.1-p22.11 Xp22.2-p22.31 Xp23 Xq27.2-q28 Xq24-q27.1 |
X:10415591-23849592 X:52987493-56318562 X:9688235-9917528 X:62853720-114517895 X:23850309-37316857 X:8784578-9687806 X:114569624-114885545 X:140429142-154494231 X:117586665-140073167 |
|
region |
breakpoint |
homologue region in human region |
|
|
cytoband |
potential Tumor associated genes |
||
|
2qC3 |
t |
2q24.3 |
2:163949907-164949906 |
|
2qE1 |
t |
11p12-p13 |
11:36116052-37116051 |
|
2qF1 |
del |
20p12.1 |
20:12702419-13702418 |
|
4qC6 |
del |
1p31.3 |
1:62401969-63401968 |
|
4qC7 |
del |
1p35.3 |
1:28974256-29974255 |
|
14qA1 |
idic |
not described |
gap |
|
15qA1 |
idic |
5p14.1 |
5:25004236-25504235 |
|
15qE1 |
t |
22q13.2 |
22:42588128-43588127 |
|
17qC |
t |
6p21.1 |
6:43581195-44581194 |
|
5qE2 |
t |
4q13 |
4:69845884-70845883 |
|
7qE1 |
t |
11q14.1 |
11:79988966-80988965 |
|
XqA4 |
dup |
Xq25 |
X:128080481-129080480 |
|
XqF5 |
dup |
Xp22.2 |
X:16106127-17106126 |
|
8qB1.1 |
t |
4q34.2 |
4:176054086-177054085 |
|
8qC2 |
del |
4q31.21 |
4:145067174-146067173 |
|
8qD3 |
del |
16q23.2 |
16:79119741-80119740 |
|
8qD1 |
del |
16q21 |
16:57292130-58292129 |
|
Table 1b: CT26 |
|||
|
region |
gain |
homologue region in human |
|
|
cytoband |
position (GRCh37/hg19) |
||
|
1qA1-qH6 |
´ +1 |
8q11.21-q12.1 8q13.1-q21.11 2q14.3-q21.1 2q11.2-q12.2 2q32.1-q32.2 13q33.1 2q32.2-qter 5q21.1-q21.2 2q14.3 2q14.1-q14.3 2q21.2-q22.1 6p12.2-p12.3 6q11.1-q13 6p11.2-p12.1 18q21.32-q22.1 Y1q11.23 1q43-q44 1q32.2-q42.13 1q32.1-q32.2 1q23.1-q32.1 1q21.1 4q26 |
8:50767106-56535248 8:67336477-76107163 2:128848553-131914911 2:97151065-106819719 2:189007277-190504466 13:103237605-103533914 2:190506076-242812118 5:98439740-102728411 2:122585948-126347698 2:114436107-122578025 2:133138389-138607743 6:49796129-52568703 6:61967179-73920868 6:56223874-58686221 18:58351903-65328593 Y:28358518-28544030 1:240253393-247125743 1:207575939-227644727 1:206075775-207534964 1:158516903-205922697 1:143881371-144095755 4:119339188-119512723 |
|
2qC3-qter |
+1 |
20q13.32-q13.33 20q11.21-13.32 20p13 20p11.21 –p13 11p11.2–p14.2 11q11.12 11p11.12 11p12.1 15q13.3–q21.2 2q13 2q13 2q11.11–q11.2 2p11.2 2q13 2q22.1-q32.1 |
20:58148222-62907435 20:29933153-58056214 20:102147-1447942 20:1736101-25606620 11:26296397-48658712 11:55080583-55323018 11:51377850-51539057 11:56082416-57753858 15:32906987-51298173 2:112973390-113650007 2:111483204-112960231 2:95642277-97040617 2:87345633-87996071 2:113723845-114137444 2:140065297-188395329 |
|
3qE2–qF2 |
+1 |
7q11.23 1q21.1-q23.1 1p11.2-p22.1 3q25.1-q26.2 4q31.3-q32.2 |
7:76282730-76575579 1:144676687-158154741 1:93905157-120696915 3:149055816-167822106 4:150966383-163096512 |
|
3qF3-qter |
+2 |
1p22.2-p31.3 4q22.3-q26 4q26-q27 |
1:68589539-89738135 4:95284699-119338945 4:119596924-120703320 |
|
5qC3-qD |
+2 |
4q12-q22.1 |
4:52689038-89000187 |
|
5qE1-qF |
+1 |
1p22.1-p22.2 4p16.3 12q24.33 12q24.11-q24.31 12q23.3-q24.11 22q11.23-q12.1 |
1:89950168-93744300 4:493106-1023731 12:132378991-133522542 12:110488793-121497537 12:108325357-110486420 22:25201765-29156283 |
|
5qF-qter |
+2 |
7p22.1-p22.3 7p15.3 7q11.23-q21.2 7q22.1-q22.3 7q36.1-q36.3 7p15.3 7q36.1 7q21.3-q22.1 7q22.1 7q11.23 7q11.21-q11.23 7p11.2 13q12.13-q13.2 12q24.31-q24.33 |
7:169204-6771649 7:22903126-23240676 7:76702246-92676514 7:102331617-105210094 7:150588373-157179154 7:22595306-22772664 7:149628652-149959756 7:97598308-99229367 7:99552841-102191754 7:72536306-76149827 7:66808098-72045725 7:56019352-56184138 13:26784894-34260463 12:121577100-132336561 |
|
6qA1–qA3 |
+2 |
7q31.1-q36.1 7p21.3-p22.1 7p21.3 |
7:112138919-149583263 7:7132996-12536829 7:92745197-97502117 |
|
6qA3-qG3 |
+1 |
1p31.3 22q11.1-q11.21 4q27 4q22.1-q22.3 10q11.21-q11.22 2p11.2-p13.3 2p11.2 3p25.2-pter 3p25.1-p25.2 3p12.3-p14.1 3q21.3 3q22.1 7p14.3-p15.3 12p11.21 12p11.21-p13.31 12p13.31-p13.33 12p13.31 12p13.33 |
1:67631910-68317098 22:17565811-18659740 4:121018693-122194687 4:89178698-95273100 10:43277986-46218167 2:68715037-87095119 2:88302422-89174373 3:61304-12897767 3:12939278-15163105 3:64017713-75322601 3:125725101-129038484 3:129094932-129632650 7:23254035-33103246 12:31424829-32537434 12:9901365-30943693 12:2903120-7695890 12:8071763-9214464 12:66113-2823666 |
|
7qA1-qD1 |
+1 |
19q13.42-q13.43 19q13.43 19q13.31-q13.33 19q12-q13.31 19q12 19q13.33-q13.41 16p13.11 11p14.3-p15.1 15q11.2 15q11.2-q13.1 15q13.1-q13.3 15q26.3 |
19:54368915-57485284 19:58523795-59089552 19:45010010-48707700 19:30093064-44860951 19:28589680-30085362 19:48800017-51921957 16:16252815-16388674 11:17403485-25251145 15:22833222-23086601 15:23914751-28586067 15:29107424-32578594 15:99080385-102265870 |
|
8qA1.1–qE2 |
+1 |
19p13.11-p13.12 19p13.2 19p13.12-p13.2 16q12.1-q22.1 16q22.1-qter 10p11.21-p11.22 1q42.13-q32.3 4q32.2-qter 4q31.1-q31.23 13q33.1-qter 13q14.3 8p22-p23.1 8p23.1 8p11.23-p12 8p11.21-p11.23 8p23.1-p23.2 8p23.2-p23.3 8p21.3-22 8p11.21 22q12.3 |
19:16163040-19774937 19:7112183-8071013 19:12745060-14683008 16:46693273-69976105 16:70109527-90110030 10:33112469-35152269 1:229404294-235324774 4:163504024-190884657 4:141251922-150892329 13:103533915-115092930 13:52435459-53211718 8:12579073-17958954 8:8108776-9640417 8:29190466-36677574 8:36716542-42505949 8:5368147-6693649 8:591286-5358752 8:18227877-20177976 8:42691750-43058925 22:33658332-35953121 |
|
10qC3-qter |
+1 |
12q23.3 22q12.3 12q13.2-q23.3 |
12:104359309-108176937 12:55351591-104351507 22:32783299-33472414 |
|
13qA3-qC3 |
+1 |
5q14.3-q15 5p15.31-p15.33 9p12-p13.1 9 q22.32-q22.33 9q21.32-q21.33 9q22.1-q22.32 9p11.2 9p13.1 5q31.1-q31.2 5q35.2-q35.3 6p22.3-p23 6p23-pter |
5:84566270-96144383 5:191425-7935441 9:65585614-65901647 9:97320957-99417669 9:86231955-90340399 9:91031851-97067712 9:43623473-43941731 9:38810965-40707569 5:134073478-137090938 5:173750964-177039611 6:15104709-20060798 6:181261-15099150 |
|
15qA1-qF3 |
+1 |
5p12-p15.31 8q22.1-q24.3 22q12.3-qter 12q12-q13.2 12p11.1 |
5:8927745-42888975 8:97446632-146158346 22:35962951-51222438 12:38607141-55072925 12:33476533-34210697 |
|
17qA1-qE5 |
+1 |
18p11.32 18p11.22-p11.32 2p16.2-p16.3 2p16.3-p23.2 2q12.2-q12.3 3p24.3-p25.1 5q21.2-q22.1 19p13.3 6p12.3-p21.2 6 p21.32-p22.1 6p21.2-p21.31 6q27 6q25.3-q27 6q25.2-q25.3 5q15-q21.1 16p13.3 5q35.1 21q22.3 19p13.12 19p13.2 |
18:861722-2534400 18:2534401-9972541 2:51709987-53282184 2:29033520-51699597 2:107383985-108798215 3:16307846-20231899 5:102759315-110063021 19:4229082-6862967 6:39266498-49681826 6:29322703-33297218 6:33359177-39058058 6:167859539-170893754 6:160103032-166797236 6:155053083-160101646 5:96202316-98405239 16:222880-3208490 5:171946752-172722349 21:43490502-45122943 19:15270296-15808207 19:8366687-8811037 |
|
18qB3-qC |
+1 |
5q22.2-q31.1 |
5:112310736-130339352 |
|
19qD1-qter |
+1 |
10q23.2-q26.11 |
10:89234113-121219507 |
|
19qA-qB |
+1 |
11q12.1-q13.3 9q21.11-q21.31 |
11:57844834-68709722 9:69086307-82777364 |
|
region |
loss |
homologue region in human |
region |
|
cytoband |
position (GRCh37/hg19) |
||
|
XqA1-qA4 |
-2 |
Xp11.23-p11.22 Xp21.1-p11.23 Xq23-q24 Xq24-q27.1 |
X:48262014-51358982 X:37364439-47520178 X:115210308-117585111 X:117586665-140073167 |
|
XqA4-qter |
-1 |
Xp22.11-p22.2 Xp11.21-p11.22 Xp22.2 Xq11.1-q23 Xp21.1-p22.11 Xp22.2-p22.31 Xp23 Xq27.2-q28 |
X:10415591-23849592 X:52987493-56318562 X:9688235-9917528 X:62853720-114517895 X:23850309-37316857 X:8784578-9687806 X:114569624-114885545 X:140429142-154494231 |
|
region |
breakpoint |
homologue region in human region |
|
|
cytoband |
potential Tumor associated genes |
||
|
1qA1 |
idic |
8q11.3 |
8:52523400-53523399 |
|
2qC3 |
t |
2q24.3 |
2:163949907-164949906 |
|
2qH2 |
t |
20q13.12 |
20:43014318-44014317 |
|
3qA1 |
idic |
8q21.13 |
8:80370572-81370571 |
|
3qE1 |
t |
3q25.32 |
3:157323645-158323644 |
|
3qF3 |
t |
1p13.3 |
1:107099268-108099267 |
|
5qA1 |
idic |
7q21.11 |
7:84124870-85124869 |
|
5qC3 |
Inv. |
4p15.2 |
4:25665078-26665077 |
|
6qA1 |
idic |
7q21.3 |
7:94036515-95036514 |
|
6qA3 |
t |
7q32 |
7:129853489-130853488 |
|
6qG3 |
t |
12p12.2 |
12:20022180-21022179 |
|
7qD1 |
t |
15q26.3 |
15:99517371-100517370 |
|
9qE1 |
del |
6q14.1 |
6:79150264-80150263 |
|
10qC1 |
t |
12q23.3 |
12:105129069-106129068 |
|
10qC3 |
t |
12q21.2 |
12:78757774-79757773 |
|
12qA1 |
idic |
2p24.2 |
2:17220394-18220393 |
|
13qA1 |
idic |
10p15.2 |
10:2609713-3609712 |
|
13A3.3 |
del |
6p24.3 |
6:6607831-7607830 |
|
13qC3 |
del |
5q14.3 |
5:89325162-90325161 |
|
14qA1 |
idic |
not described |
gap |
|
15A1 |
idic |
5p14.1 |
5:24896354-25896353 |
|
15qF1 |
t |
12q12 |
12:45812915-46812914 |
|
17qA1 |
idic |
6q25.3 |
6:156599064-157599063 |
|
18qB3 |
dup |
5q31.3 |
5:141471744-142471743 |
|
19qB |
del |
9q21.13 |
9:74636718-75636717 |
|
19qD |
t |
10q24.32 |
10:104178072-105178071 |
|
XqA4 |
t |
Xq25 |
X:128080481-129080480 |
|
XqA5 |
t |
Xq25 |
X:128080481-129080480 |
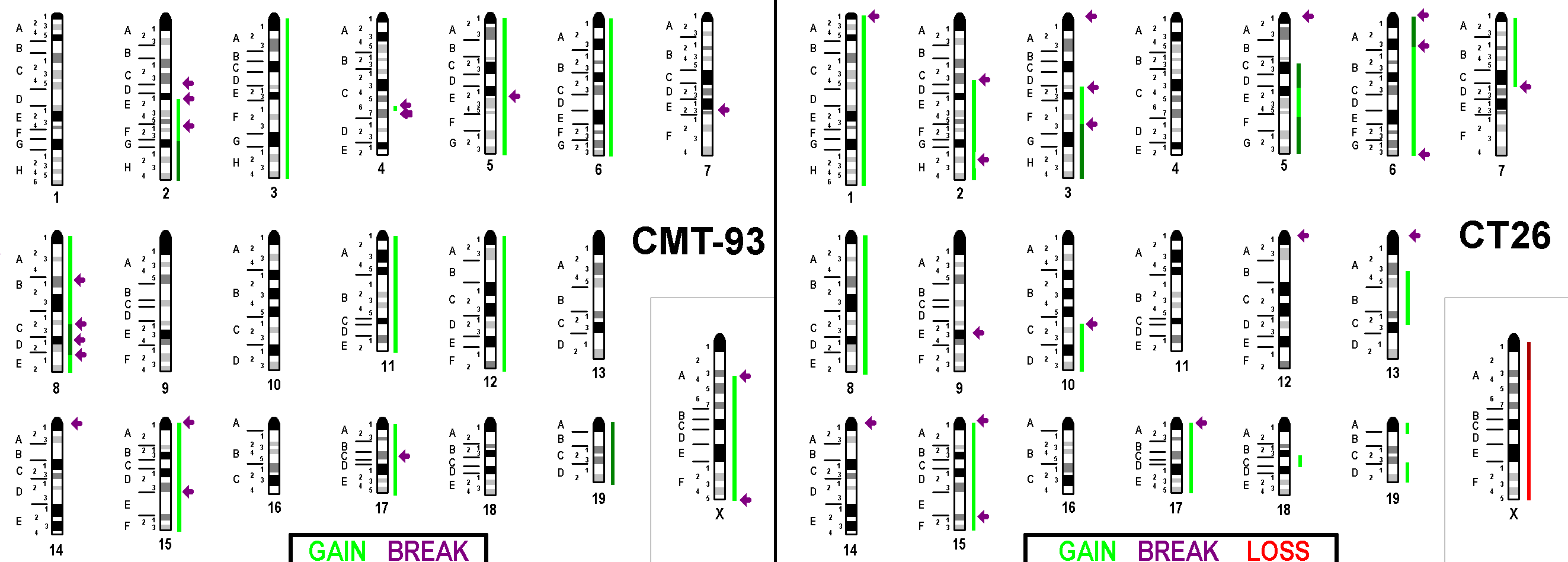
Figure 3 Copy number variations detected in CMT-93 and CT26 are summarized here with respect to a diploid-basic karyotype. Gains are depicted as green bars (one more copy = light green; two more copies = dark green), loss of one copy is depicted as a red bar and loss of two copies is depicted as a dark-red bar. Breaks are registered here as arrows. The Y-chromosome was excluded from analyses for CMT-93.

Figure 4 Imbalances present in CMT-93 and CT26 (Figure 3) were ‘translated’ to the corresponding homologous regions in human. As before, gains are depicted as green bars (one more copy = light green; two more copies = dark green), loss of one copy is depicted as a red bar and loss of two copies is depicted as a dark-red bar. Breaks are registered here as arrows. Y-chromosome was excluded from analyses for CMT-93.
According to the literature, gains of copy numbers as seen in CMT-93 and CT26 can also be observed in human CRCs. We found six regions in common with primary tumors, however, and nine regions in common with CRC metastases (Table 2).
Interestingly, regions with amplifications in the two studied cell lines harbor tumor suppressors rather than oncogenes (Table 3).
Table 2 The gain if copy numbers, as seen in CMT-93 and CT26, are compared to the gain of copy numbers observed in human CRC acc. to [17]. x = complete overlap, (x) = partial overlap.
|
|
Amplifications in >15 % of the human CRCs |
CMT-93 |
CT26 |
|
|
primary tumor |
liver metastasis |
|||
|
1q21-q41 |
- |
+ |
(x) |
x |
|
6p11-p25 |
- |
+ |
(x) |
(x) |
|
6q12 |
- |
+ |
|
x |
|
6q21 |
- |
+ |
|
|
|
7p22-q36 |
+ |
+ |
(x) |
(x) |
|
8p11-p12 |
- |
+ |
x |
x |
|
8q11-q24 |
+ |
+ |
(x) |
x |
|
11p15 |
- |
+ |
|
x |
|
11q13 |
- |
+ |
x |
x |
|
12p13-q13 |
- |
+ |
(x) |
(x) |
|
13q11-q34 |
+ |
+ |
(x) |
(x) |
|
16p13-q24 |
- |
+ |
(x) |
(x) |
|
17q11-q25 |
- |
+ |
x |
|
|
19p13 |
- |
+ |
x |
x |
|
19q13 |
- |
+ |
x |
x |
|
20p13-q13 |
+ |
+ |
x |
x |
|
Xp22 |
- |
+ |
|
|
|
Xp21-q25 |
+ |
+ |
(x) |
|
|
Xq26-q28 |
- |
+ |
(x) |
|
|
overall |
6 (9) |
9 (6) |
||
Table 3 Tumor suppressor genes of importance in CRC acc. to [17] and their involvement in gains of copy numbers in the two studied cell lines.
|
tumor suppressor genes |
gene loci |
deletion or amplification |
||
|
CRC in >15 % |
CMT-93 |
CT26 |
||
|
PI3CA |
3q26.32 |
no changes observed (nco) |
amplified |
amplified |
|
APC |
5q22.2 |
deleted |
nco |
amplified |
|
BRAF |
7q34 |
amplified |
amplified |
amplified |
|
CDKN2A |
9p21.3 |
deleted in liver metastasis |
nco |
nco |
|
TP53 |
17p13.1 |
deleted |
amplified |
nco |
In Table 4 breakpoints observed in the two cell lines studied were compared to those known to appear in human CRCs. In CMT-93, five, and in CT26, eight, breakpoints were in concordance with human CRC-associated breakpoints.
Table 4 Breakpoints in CMT-93 and CT26 compared to the 50 most frequently observed acquired breaks in human CRCs [13].
|
breakpoint acc. to human genome |
breakpoint observed in |
Breakpoint in human CRCs |
|
|
CMT-93 |
CT26 |
||
|
1p35.3 |
+ |
- |
(x) |
|
1p31.3 |
+ |
- |
(x) |
|
1p13.3 |
- |
+ |
x |
|
2p24.2* |
+ |
+ |
- |
|
2q24.3 |
+ |
+ |
- |
|
3p14.2* |
+ |
+ |
- |
|
3q25.32 |
- |
+ |
- |
|
4p16.3-q22.1 |
- |
+ |
- |
|
4p12 |
+ |
+ |
- |
|
4q34.2 |
+ |
- |
- |
|
5p14.1* |
+ |
+ |
- |
|
5q14.3 |
- |
+ |
- |
|
5q23.2 |
- |
+ |
- |
|
6p24.3 |
- |
+ |
- |
|
6p21.1 |
+ |
- |
- |
|
6q14.1 |
- |
+ |
(x) |
|
6q25.3* |
- |
+ |
- |
|
7q21.11* |
- |
+ |
- |
|
7q21.3* |
- |
+ |
- |
|
7q34 |
- |
+ |
- |
|
8q11.3* |
- |
+ |
- |
|
8q21.13* |
- |
+ |
- |
|
9q21.13 |
- |
+ |
(x) |
|
10p15.2* |
- |
+ |
- |
|
10q24.32 |
- |
+ |
x |
|
11p15.5 |
+ |
- |
- |
|
11p15.4 |
+ |
- |
- |
|
12p12.2 |
- |
+ |
- |
|
12q12 |
- |
+ |
(x) |
|
12q23.3 |
- |
+ |
(x) |
|
15q25.3 |
- |
+ |
- |
|
16q21 |
+ |
- |
- |
|
16q22.1 |
+ |
- |
- |
|
19p13.13 |
+ |
- |
x |
|
20p12.1 |
+ |
- |
(x) |
|
20q13.12 |
- |
+ |
- |
|
22q13.2 |
+ |
- |
- |
|
Xp22.2 |
+ |
- |
+ |
|
Xq25 |
+ |
+ |
- |
|
Xq27.1-q27.2 |
- |
+ |
- |
* = breaks in centromere-region of murine genome, x = concordance on molecular level, (x) = concordance on molecular cytogenetic level.
Finally, we must state that chromothripsis was found in about 1.0% of CMT-93 metaphases, but it was not observed for CT26.
4. Discussion
Overall, murine-tumor cell lines are under-characterized concerning their (cyto) genetic constitution; this also holds true for the CRC cell lines, CMT-93 and CT26, used for this study. Accordingly, the karyotypes of CMT-93 and CT26 were characterized in detail based on mcb and aCGH.
Molecular cytogenetics (mFISH and mcb) are intended to provide a comprehensive cytogenetic description, recording ploidy, numerical and structural aberrations, and clonal and nonclonal changes, and localizing the associated breakpoints. Compared to classical banding techniques, mFISH allows a simple and unambiguous identification of all mouse chromosomes, and provides an overview of all existing gross numerical and structural chromosome aberrations [19]. In common with other whole-chromosome painting methods, mFISH-only is not suitable for identifying intrachromosomal rearrangements such as duplications, deletions, or inversions [20]. Thus, many of the unclear findings that mFISH provided (e.g., for small translocations and deletions) can be resolved by the FISH-banding approaches, as those applied in this study, such as mcb; furthermore, aCGH has successfully been applied for better characterization of breakpoints in cases of unbalanced rearrangements.
Overall, both cell lines showed clonal changes in common and in general had a tendency for ongoing karyotypic evolution [4,21]. Additionally, it must be mentioned that the studied cell lines were established about 40 years ago and have been passaged many times since then. Thus, the development of subclones is a usual finding [9,10,11], as karyotype evolution is naturally linked with solid tumors [4]. Furthermore, a tendency towards polyploidization was observed in these cell lines, which is also present in other murine-tumor cell lines [22]. This was suggested to be due to telomere-driven tetraploidization in the context of tumor progression [23], and/or cell culture-related factors such as trypsin treatment, increasing number of cell-culture passages, and oxygen exposure [24]. Overall, the karyotype evolution of cell lines is well known; however, only few labs consider re-evaluating the genomic profile of these cell lines that have been used for many years.
In addition, in CMT-93 and CT26 (except for the gonosomes) we observed only gains and no loss of copy numbers. This is in contrast to the situation in human CRCs, which also show a number of typical deletions (Table 3) [17]. However, this is in concordance with molecular studies performed, especially in CMT-93 [1,2,25]. Interestingly, in both cell lines, the not-yet-observed derivative chromosomes was detected, which either have a centromere-near deletion with a centromeric and a more distal breakpoint, or a neocentromere after the loss of the proximal chromosomal part. There are neocentromeres reported in malignancies [26]; still, such chromosomes are considered rather instable [27].
Chromothripsis is nowadays discussed to be a common phenomenon in cancer. This is also valid for, and has been observed in, primary and metastatic CRCs [18,28]. Chromothripsis has also been observed previously in other murine cell lines [11], in this study for CMT-93, in a small subset of the cells. Possibly, this could explain the slightly lower polyploidy rate of CMT-93 compared to CT26 and would be consistent with an advanced tumor stage.
The present study was focused on the detection and characterization of gross chromosomal rearrangements in the two cell lines we studied. Based on this, still existing gaps in the profiling of CMT-93 and CT26 that were not covered here, like point mutations, RNA- or proteomic features, can be performed in future.
Overall, it could be shown that, based on chromosomal imbalances (Table 2), the cell lines utilized in our study have major chromosomal aberrations as both primary and metastatic CRCs. Still, they clearly resemble advanced types of metastatic CRCs. For copy number changes of tumorsuppressorgenes, as listed in Table 3 and for chromosomal breakpoints (Table 4), the concordance is less expressed, but still partially present. Of special interest may be the fact that eight of the breakpoints listed in Table 4 are located in murine centromeres, which are heterochromatic and cannot be attributed to human homologous regions. Similar heterochromatic breakpoints are rarely reported for human cancer [29]. On top of that, both cell lines were induced initially by chemicals, which may also lead to different breakpoints than in human CRCs. Finally, the fact that the mouse line diverged from the human line about 200 million years ago may explain some of the differences we observed relating to the tumor-associated breakpoints.
5. Conclusions
In summary, both cell lines show major similarities to genetic peculiarities, as known from human metastatic CRCs. Thus, they are model systems for advanced and not primary CRCs; or, as stated for CT26 already [14], they are models for aggressive, undifferentiated, and refractory human CRCs.
Author Contributions
NK did the cell line cultivation and preparation of chromosomes and DNA; HR and SSHA performed the practical molecular cytogenetic work; MR did the aCGH (practical work and evaluation); TL drafted the study and finalized the paper together with the other authors
Funding
Supported by grant # 2013.032.1 of the Wilhelm Sander-Stiftung.
Competing Interests
The authors have declared that no competing interests exist.
References
- Wlochowitz D, Haubrock M, Arackal J, Bleckmann A, Wolff A, Beissbarth T, et al. Computational identification of key regulators in two different colorectal cancer cell lines. Front Genet. 2016; 7: 42. [CrossRef]
- Marmol I, Sanchez-de-Diego C, Pradilla Dieste A, Cerrada E, Rodriguez Yoldi MJ. Colorectal carcinoma: A general overview and future perspectives in colorectal cancer. Int J Mol Sci. 2017; 18: 197. [CrossRef]
- Tong Y, Yang W, Koeffler HP. Mouse models of colorectal cancer. Chin J Cancer. 2011; 30: 450-462. [CrossRef]
- Ye CJ, Regan S, Liu G, Alemara S, Heng HH. Understanding aneuploidy in cancer through the lens of system inheritance, fuzzy inheritance and emergence of new genome systems. Mol Cytogenet. 2018; 11: 31. [CrossRef]
- Gordon J, Fischer-Cartlidge E, Barton-Burke M. The big 3: An updated overview of colorectal, breast, and prostate cancers. Nurs Clin North Am. 2017; 52: 27-52. [CrossRef]
- Eppig J, Richardson J, Kadin J, Smith C, Blake J, Bult C. Mouse genome database: From sequence to phenotypes and disease models. Genesis. 2015; 53: 458-473. [CrossRef]
- Day C, Merlino G, Van Dyke T. Preclinical mouse cancer models: A maze of opportunities and challenges. Cell. 2015; 163: 39-53. [CrossRef]
- Carver BS, Pandolfi PP. Mouse modeling in oncologic preclinical and translational research. Clin Cancer Res. 2006; 12: 5305-5311. [CrossRef]
- Leibiger C, Kosyakova N, Mkrtchyan H, Glei M, Trifonov V, Liehr T. First molecular cytogenetic high resolution characterization of the NIH 3T3 cell line by murine multicolor banding. J Histochem Cytochem. 2013; 61: 306-312. [CrossRef]
- Kubicova E, Trifonov V, Borovecki F, Liehr T, Rincic M, Kosyakova N, et al. First molecular cytogenetic characterization of murine malignant mesothelioma cell line AE17 and in silico translation to the human genome. Curr Bioinform. 2017; 12: 11-18. [CrossRef]
- Guja K, Liehr T, Rincic M, Kosyakova N, Hussein Azawi SS. Molecular cytogenetic characterization identified the murine B-cell lymphoma cell line A-20 as a model for sporadic Burkitt's lymphoma. J Histochem Cytochem. 2017; 65: 669-677. [CrossRef]
- Golovko D, Kedrin D, Yilmaz OH, Roper J. Colorectal cancer models for novel drug discovery. Expert Opin Drug Discov. 2015; 10: 1217-1229. [CrossRef]
- Knutsen T, Padilla-Nash HM, Wangsa D, Barenboim-Stapleton L, Camps J, McNeil N, et al. Definitive molecular cytogenetic characterization of 15 colorectal cancer cell lines. Genes Chromosomes Cancer. 2010; 49: 204-223.
- Castle JC, Loewer M, Boegel S, de Graaf J, Bender C, Tadmor AD, et al. Immunomic, genomic and transcriptomic characterization of CT26 colorectal carcinoma. BMC Genomics. 2014; 15: 190. [CrossRef]
- Claussen U, Michel S, Mühlig P, Westermann M, Grummt UW, Kromeyer-Hauschild K, et al. Demystifying chromosome preparation and the implications for the concept of chromosome condensation during mitosis. Cytogenet Genome Res. 2002; 98: 136-146. [CrossRef]
- Liehr T, Starke H, Heller A, Kosyakova N, Mrasek K, Gross M, et al. Multicolor fluorescence in situ hybridization (FISH) applied to FISH-banding. Cytogenet Genome Res. 2006; 114: 240-244. [CrossRef]
- Diep C, Kleivi K, Ribeiro F, Teixeira M, Lindgjaerde O, Lothe R. The order of genetic events associated with colorectal cancer progression inferred from meta-analysis of copy number changes. Genes Chromosomes Cancer. 2006; 45: 31-41. [CrossRef]
- Skuja E, Kalniete D, Nakazawa-Miklasevica M, Daneberga Z, Abolins A, Purkalne G, et al. Chromothripsis and progression-free survival in metastatic colorectal cancer. Mol Clin Oncol. 2017; 6: 182-186. [CrossRef]
- Schröck E, Zschieschang P, O'Brien P, Helmrich A, Hardt T, Matthaei A, et al. Spectral karyotyping of human, mouse, rat and ape chromosomes--applications for genetic diagnostics and research. Cytogenet Genome Res. 2006, 114: 199-221. [CrossRef]
- Kearney L. Multiplex-FISH (M-FISH): technique, developments and applications. Cytogenet Genome Res. 2006; 114: 189-198. [CrossRef]
- Bloomfield M, Duesberg P. Is cancer progression caused by gradual or simultaneous acquisitions of new chromosomes?. Mol Cytogenet. 2018; 11: 4. [CrossRef]
- Levan A, Biesele JJ. Role of chromosomes in cancerogenesis, as studied in serial tissue culture of mammalian cells. Ann N Y Acad Sci. 1958, 71: 1022-1053. [CrossRef]
- Davoli T, de Lange T. Telomere-driven tetraploidization occurs in human cells undergoing crisis and promotes transformation of mouse cells. Cancer Cell. 2012; 21: 765-776. [CrossRef]
- Mastromonaco GF, Perrault SD, Betts DH, King WA. Role of chromosome stability and telomere length in the production of viable cell lines for somatic cell nuclear transfer. BMC Dev Biol. 2006; 6: 41. [CrossRef]
- Guda K, Upender MB, Belinsky G, Flynn C, Nakanishi M, Marino JN, et al. Carcinogen-induced colon tumors in mice are chromosomally stable and are characterized by low-level microsatellite instability. Oncogene. 2004; 23: 3813-3821. [CrossRef]
- de Figueiredo AF, Mkrtchyan H, Liehr T, Soares Ventura EM, de Jesus Marques-Salles T, Santos N, et al. A case of childhood acute myeloid leukemia AML (M5) with a neocentric chromosome neo(1)(qter-->q23 approximately 24::q23 approximately 24-->q43-->neo-->q43-->qter) and tetrasomy of chromosomes 8 and 21. Cancer Genet Cytogenet. 2009; 193: 123-126. [CrossRef]
- Burrack LS, Hutton HF, Matter KJ., Clancey SA, Liachko I, Plemmons AE, et al. Neocentromeres provide chromosome segregation accuracy and centromere clustering to multiple loci along a Candida albicans chromosome. PLOSGenetics. 2016; 12: e1006317. [CrossRef]
- Kloosterman WP, Hoogstraat M, Paling O, Tavakoli-Yaraki M, Renkens I, Vermaat JS, et al. Chromothripsis is a common mechanism driving genomic rearrangements in primary and metastatic colorectal cancer. Genome Biol. 2011; 12: R103. [CrossRef]
- Backsch C, Pauly B, Liesenfeld M, Scheungraber C, Gajda M, Mrasek K, et al. Two novel unbalanced whole arm translocations are frequently detected in cervical squamous cell carcinoma. Cancer Genet. 2011; 204: 646-653 [CrossRef]
